
Abstract
The present investigation demonstrates establishment of United Nations Environment Programme (UNEP) priority Persistent Organic Pollutants (POPs) using high-resolution gas chromatography–high-resolution mass spectrometry. Particularly, POPs analytical methods were established using native and 13C-labeled internal standards of HCHs, HCB, cyclodienes, chlordanes, DDTs, mirex, dioxin-like PCBs, PCDDs, and PCDFs by isotope dilution technique. The relative response factor for 6-point calibration curve native standards (18 replicate analysis) were in the ranges of 0.93–1.43 with relative standard deviation ranges from 1.68 to 4.96%. Instrument detection limit and instrument quantification limit was established for various POPs at femtograms. Concentrations of UNEP-POPs were measured in liver (n = 10) and egg (n = 10) of great cormorants and their major diet, gizzard shad (n = 2), collected in and around Tokyo, Japan. DDTs (ranges in liver and egg, respectively) were predominant accumulants (9800–310,000 and 9600–73,000) followed by dioxin-like PCBs (4500–69,000 and 7900–150,000), chlordanes (2600–16,000 and 700–4,800), cyclodienes (650–4600 and <1–1000), HCB (680–2800 and 180–590), HCHs (230–1800 and 120–490), PCDD/DFs (3.2–27 and 1.7–5.7) on nanogram per gram lipid basis. Concentrations (ranges) of POPs in gizzard shad were in the following order: DDTs (3900–16,000), chlordanes (3400–14,000), cyclodienes (340–1300), HCB (110–480), and HCHs (140–360) on nanogram per gram lipid basis.
Similar content being viewed by others
Continued monitoring of organochlorine pesticides (OCPs) is necessary due to their greater recalcitrant nature, biomagnification in the food chain, and long-term reproductive/health effects in wildlife and humans. Among them, DDTs ([dichloro 2,2′-bis(p-chlorophenyl)-1,1,1-trichloroethane] o,p′- and p,p′-compounds of DDT, DDE, and DDD), HCHs (α,β,γ,δ-hexachlorocyclohexane isomers), chlordanes (cis/trans-chlordane, cis/trans-nonachlor, oxychlordane, heptachlor, and heptachlor epoxide), hexachlorobenzene (HCB), cyclodienes (aldrin, dieldrin, endrin), toxaphene, and mirex are of significant importance (UNEP Chemicals, GMN 2001, 2002). These groups of compounds have been identified as priority environmental contaminants posing a significant impact to human and wildlife.
Similarly, polychlorinated diaromatic hydrocarbons (PCDHs), such as polychlorinated biphenyls (PCBs), are a group of synthetic chemicals that were used between the 1880s and the 1970s. Polychlorinated-dibenzo-p-dioxins (PCDDs), and -dibenzofurans (PCDFs), unlike PCBs, have not been purposely manufactured, but rather are present as impurities associated with chlorophenols [for example, pentachlorophenol (PCP)], in herbicides, such as 2,4,5-trichlorophenoxyaceticacid (2,4,5-T) and chloronitrophen (CNP) applied to paddy fields in Japan (Masunaga et al. 2001a,b). While PCBs had been used in various industrial materials, such as transformers, capacitors, and noncarbon copying paper, PCDD/DFs are also formed by photochemical and thermal reactions during and after municipal solid waste incinerators (MSWI) and industrial waste incinerators (IWI) (Baker and Hites, 2000; Iino et al. 2000; Hosomi et al. 2003). The PCDHs also listed as United Nations Environment Programmes (UNEPs) priority compounds (persistent organic pollutants [POPs]; a dirty dozen) with similar reasons described for OCPs, which showed a variety of toxic effects in humans and wildlife (Birnbaum and Tuomisto 2000; Feeley and Brouwer 2000; Sweeney and Mocarelli 2000; van den Berg et al. 2000).
The great cormorant (Phalacrocorax carbo) is a major predator in the freshwater food chain, and it was therefore expected that they would be highly contaminated by POPs (Tillitt et al. 1992; Sanderson et al. 1994; van den Berg et al. 1994, 1995; Boudewijn and Dirksen 1995; Williams et al. 1995; Larson et al. 1996; Meadows et al. 1996; Guruge et al. 1997; Powell et al. 1997; Custer et al. 1999, 2001; Kannan et al. 2001). In addition, great cormorants accumulated greater amounts of chlorinated organohalogens when compared to other birds and wildlife collected from Japan (Guruge et al. 1997, 2000; Iseki et al. 2001a,b; Kang et al. 2002; Senthil Kumar et al. 2002a, 2003a). This could be a reason for recently observed high chick and juvenile mortality in the colonies in and around Tokyo, Japan (Guruge et al. 2000; Iseki et al. 2001a,b).
In order to provide precise analytical data, establishment of UNEP-POPs with updated analytical procedure is essential. For example, high-resolution gas chromatography–high-resolution mass spectrometry (HRGC-HRMS) gives accurate quantification (Yamashita et al. 2002; Yasuda et al. 2003; Takasuga et al. 2003). In addition, there are several sensitive species whose habitat in pristine environments could be affected even at trace contaminant levels. The isotope-labeled pesticides standards and isotope dilution method support HRGC-HRMS to provide precise analysis of contaminants as similarly labeled PCDD/DFs do. Consequently in this study, we established a high-quality method of analyzing 11 among 12 priority UNEP POPs. Analysis of toxaphene based on electron ionization (EI: the current method) was not feasible; nevertheless, electron capture negative ionization (ECNI) was found relevant for toxaphene analysis described elsewhere (Matsukami et al. 2004) and thus except for toxaphene, other UNEP POPs could be determined using our present study method. Consequently, we also analyzed most of UNEP POPs (except toxaphene and mirex) in liver and eggs of great cormorants and their major diet, gizzard shad (fish), collected in and around Tokyo, Japan. Eventually, some background information on OCPs concentrations and toxic equivalency (TEQ) of dioxin-like PCBs and PCDD/DFs were calculated and discussed.
Materials and Methods
Chemicals
All organic solvents used for extraction and cleanup were of pesticide residue analysis grade, and were purchased from the Kanto Chemicals Co., Inc. (Tokyo, Japan) and Wako Pure Chemicals Industries, Ltd. (Osaka, Japan). 13C12-labeled and unlabeled native standards of OCPs, PCDD/DFs and PCBs were purchased from Wellington Laboratories (Ontario, Canada) and Cambridge Isotope Laboratories (Massachusetts, USA). All the original standards were prepared with different solvents (e.g., acetone, decane, methanol, acetonitrile, etc.).
The native (22 individual compounds) and 13C-labeled standards (20 individual compounds) of OCPs were used for preliminary calibration purpose. Uniform concentrations of native standards (22-mix) were mixed together, and its relative response factor (RRF) was performed (Table 1) using six different native standard dilutions (0.4, 2, 10, 40, 200, and 1000 pg/μL) after 18 replicate analysis. The 13C-labeled standards and syringe spike (injection recovery standard) concentration was set as 40 pg/μL. All these standard mixtures were analyzed in HRGC-HRMS. In order to understand sensitivity of HRGC-HRMS and HRGC-LRMS (low-resolution mass spectrometer), in addition, approximately 200 fg of the OCP standard mixture was prepared and analyzed in both with LRMS resolution of less than 1500. The detailed HRGC-HRMS parameters are discussed in later sections.
Samples
Great cormorants (Phalacrocorax carbo) were collected among birds shot under license from the Japanese Government (in order to examine the diet composition: also see Watanabe et al. 2004) in 2000 along the Sagami River in Kanagawa Prefecture of Japan, which is approximately 40 km southwest of Metropolitan Tokyo. Upon collection, the birds were dissected and the liver was separated and stored in chemically clean vials, transported to the laboratory with dry ice, and stored at −30°C until analyzed. The eggs were collected from Odaiba, in the Minato ward of Tokyo Bay in 1998, which is the site of a substantial cormorant colony roost in Japan. After washing the surface of the eggshells with an ordinary detergent and acetone, the ingredients were transferred individually to acetone/hexane-washed glass bottles and were sealed and stored in a freezer. Great cormorant major diet such as gizzard shad (Konosirus punctatus) a fish was collected from Tokyo Bay in 1998. The fish was whole body homogenated for chemical analysis and packed in chemically clean polythene bags and stored in freezer. The sampling details and few pretreatment process details are summarized elsewhere Watanabe et al. (2004).
Extraction and Cleanup
Prior to extraction, all the samples were freeze-dried for 48 hours and their initial and final weight was measured before and after freeze-dry, respectively, and then powdered in mortar and pestle. The sampling spikes such as 13C6-δ-HCH, 13C10-cis-nonachlor, and 13C12-o,p′-DDE was spiked in known amount (approximately, 5 g fresh liver, 20 g fresh fish, and fresh egg) of samples. The spiked liver samples were extracted with accelerated solvent extractor (ASE) with dichloromethane (DCM) for 30 min with ASE standard operating procedure (SOP), whereas spiked egg and fish were extracted with DCM using Soxhlet extractor for 16 hours. All the extracted solvents were rotary evaporated to 10 mL and lipid weight was measured using 1 mL of extracts. From the 9 mL of remaining extracts, 2 mL was used for OCP analysis by further spiking 13C internal standards listed in Table 1. The overall analytical procedure of UNEP POPs is summarized in Figure 1. Briefly, after lipid measurement, the extracts were resolved for OCPs and dioxin-like PCBs and PCDD/DFs analysis separately. For OCP analysis, in the extracts, the known amount of 13C internal standards of OCPs (Table 1) were spiked, passed through the activated florisil column, and collected with 150 mL of 100% DCM in hexane. The eluted solvent was then rotary evaporated to 2 mL and subjected to dimethyl sulfoxide (DMSO)–hexane (1:1 v/v) partitioning in order to remove the remaining lipid content. After DMSO–hexane partitioning, the samples were further fractionated with activated florisil. Fraction A with 20% DCM in hexane contained HCH isomers, HCB, aldrin, heptachlor, chlordane compounds, mirex, and DDT and its metabolites, whereas fraction B collected with 100% DCM contained dieldrin, endrin, and heptachlor epoxide. Both fractions were syringe spiked (TeCB-70), or injection recovery spike was included prior to HRGC-HRMS analysis.

The schematic diagram showing analytical procedure of UNEP POPs.
The cleanup for dioxin-like PCBs and PCDD/DFs is also summarized in Figure 1. Briefly, in the lipid-determined extracts, the known amount of 13C12-labeled 2,3,7,8-PCDD/DFs and dioxin-like PCBs internal standards were spiked, passed through multilayer silicagel [anhydrous Na2SO4 (1 g), silica (1 g), 10% (w/w) AgNO3-silica (1 g), silica (0.5 g), 22% (w/w) H2SO4-silica (3 g), 44% (w/w) H2SO4-silica (5 g), silica (0.5 g), 2% (w/w) KOH-silica (0.5 g), silica (0.5 g), and anhydrous Na2SO4 (1 g)] column chromatography, and collected with hexane as mobile phase. The multilayer-fractionated samples were further fractionated with silica gel–dispersed carbon column and eluted with 25% DCM in hexane as a prefraction that contained mono- and di-ortho PCBs. The post fraction was done with toluene that contained non-ortho PCBs and PCDD/DFs. For dioxin-like PCBs the syringe spike (PeCB-111) was included, while 1,2,3,4-TeCDD was included as syringe spike for PCDD/DFs prior to HRGC-HRMS analysis.
Quantification and Identification
Quantification and identification of OCPs were performed with the Hewlett-Packard HP-6890 Series Micromass Autospec Ultima Series HRGC-HRMS system. Both DB-5MS and DB-17HT columns were used for OCP analysis. An autosampler (GC System Injector, Hewlet-Packard) was used for injection (2 μL, on-column). The HRGC-HRMS program for OCPs had a resolution of more than 10,000 with selected ion monitoring (SIM) as summarized in Table 2. The column temperature was 120°C (1 min) - 20°C/min-160°C (0 min) - 3°C/min - 220°C (0 min) -10°C/min - 300°C (1 min). The on-column temperature was 120°C (0.1 min) - 100°C/min - 300°C (15 min). The interface temperature was 300°C. Helium was used as carrier gas at 1 mL/min flow rate.
Quantification and identification of dioxin-like PCBs and 2,3,7,8-substituted congeners of PCDD/DFs were performed using a Hewlett-Packard HP-6890 Series Micromass Autospec Ultima Series HRGC-HRMS system. Both DB-5MS (J&W Scientific; 0.25 mm i.d. × 60-m length for PCBs) and a DB-17HT (J&W Scientific; 0.32 mm i.d. × 30-m length) columns were used for PCDD/DFs. On the other hand, a DB-5MS column was utilized for the analysis of dioxin-like PCBs. An autosampler (GC System Injector, Hewlett-Packard) was used for injection (2 μL, on-column). The temperature programs used for PCDD/DF determination are as follows: 130°C for 1 min, 20°C/min to 200°C for 0 min, hold for 2 min, 3°C/min to 250°C for 0 min, 5°C/min, to 300°C for 3 min (DB-17 HT). In case of dioxin-like PCBs, the temperature programs used were as follows: 150°C for 1 min, 20°C/min to 185°C for 1 min and hold for 2 min, 3°C/min to 245°C for 0 min, 6°C/min to 290°C.... The temperatures of the injector and the ion source were 280°C and 250°C, respectively. The interface temperature was set at the maximum value of each temperature program. The carrier gas was helium and the electron impact ionization energy was 40 eV. The SIM mode was used and the resolution was kept higher than 10,000 (>5% valley). A blank run was conducted during each batch consisting of five samples. However, blanks did not contain detectable limits of analytes.
Results and Discussion
General Discussions on Merits of Method
All the 22 mixture native standards RRF established in this study were in the range of 1.08–1.24 (HCH isomers), 1.03 (HCB), 1.17–1.24 (cyclodienes), 0.93–1.18 (chlordanes), 1.06–1.43 (DDTs and its metabolites), and 1.03 (mirex) with the percent standard deviation on the order of the scientifically acceptable range, 1.68 to 4.96% (Table 1). Unlabeled and 13C-labeled standard OCPs were monitored at different mass ratio m/z (Table 2) with m/z between 181 and 409. DB-5MS and DB-17HT columns were used for OCPs analysis, whereas DB-5MS showed interference (same retention time) of heptachlor epoxide and oxychlordane and cis-nonachlor and p,p′-DDD and therefore only DB-17HT columns (30 m × 0.32 mm i.d. [0.15 μm]) were used for OCP analysis without any interference and well-resolved individual peaks.
The instrument detection limits (IDL), and instrument quantification limit (IQL) was calculated by 3-times and 10-times multiplication, respectively, of the standard deviation (STDEV) of six replicate analysis of the OCP native standard (Table 3). The IDL and IQL were classified in the decreasing order: HCB followed by o,p′-DDT, α-HCH, p,p′-DDD, p,p′-DDT, o,p′-DDE, cis-nonachlor, trans-chlordane, γ-HCH, o,p′-DDD, δ-HCH, heptachlor, mirex, p,p′-DDE, cis-chlordane, aldrin, β-HCH, heptachlor epoxide, dieldrin, trans-nonachlor, oxychlordane, and endrin as far as obtained concentration is concerned (Table 3).
Furthermore, quantification of UNEP POPs was also conducted using HRGC-LRMS for comparison purposes. The analysis of all OCPs in HRGC-HRMS and HRGC-LRMS showed magnitude of difference with detection limits. The former showed detection of femtograms, whereas the latter showed detection of picograms and therefore trace concentrations of UNEP POPs can be monitored using the former (Table 3). Based on these results, it can be justified that HRGC-HRMS methods are very useful for samples collected from remote marine locations because of more selectivity. Particularly, the method is more useful for air samples collected from pristine environments such as Arctic and Antarctic regions in which the animals that inhabit them are considered to be very sensitive even to trace concentrations of pollutants.
UNEP POPs
Recoveries
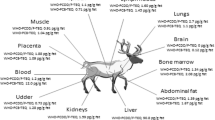
The percentage recoveries of UNEP POPs that spiked into cormorant liver, egg, and their diet (gizzard shad) and passed through the whole analytical procedure are listed in Table 4. Percentage recoveries for sampling spikes (δ-HCH, cis-nonachlor and o,p′-DDE) were greater than procedural (internal) standards. Perhaps the sampling standard (3-mix) and cleanup standards (19-mix) were prepared at different periods, which would have influenced recoveries. In addition, original standards that were purchased were synthesized with different solvents that considered to possible explanation. Nevertheless, there was not much matrix effect on recoveries because liver, egg, and gizzard shad showed almost similar recoveries. Percentage recovery was not analyzed for gizzard shad because analysis of dioxin-like PCBs and PCDD/DFs were not conducted for these samples. When comparing liver and egg recoveries, PCDD/DFs showed greater recovery followed by dioxin-like PCBs, chlordanes, DDTs, cyclodienes, HCHs, and HCB. Degradation of HCB, heptachlor, and HCHs during nitrogen purge and rotary evaporation was assumed to be the most critical step for the decreased recovery, whereas florisil fractionation was assumed to be the most critical step for the decreased endrin recovery. Concentrations of all compounds were corrected to recovery RRF of 6-calibration curve. The concentrations indicated in this study were expressed as nanograms per gram lipid basis (OCPs and dioxin-like PCBs) and picograms per gram lipid basis (PCDD/DFs and TEQ) unless otherwise specified. The values were rounded to two significant numbers.
HCHs
The average concentrations of HCHs in cormorant liver, egg, and gizzard shad (fish) are summarized in Table 5. The contribution of HCHs to total UNEP POPs load is 0.65, 0.31, and 1.28% in liver, egg, and fish, respectively. In all the sample matrices, β-HCH was the predominant isomer to the total HCHs. Predominance of β-HCH could be explained as greater stability of this isomer to the enzymatic degradation in animal body as observed in several biota samples (Senthil Kumar et al. 2001a). Guruge et al. (1997) reported similar HCH composition in liver of cormorants collected from Lake Biwa and Shinobazu Pond in Tokyo, Japan. HCHs were used in Japan in between February 24, 1949 and December 30, 1971 and then banned (Uemura et al. 2002). Occurrence of HCHs in livers and eggs of cormorants and fish possibly resulted from the prolonged use of these chemicals in southern Asian countries such as India and China. Atmospheric transport of HCHs is probably a plausible explanation for the trans-boundary contamination and continued occurrence in wildlife in Japan (Senthil Kumar et al. 2001a). The HCHs were not included in UNEP’s priority POPs list, although they have long-range transport properties and bioaccumulation potential in the food web.
HCB
Mean concentrations of HCB in cormorant liver, egg, and fish, shown in Table 5, which are 1.1% (liver), 0.53% (egg), and 1.5% (fish) to the total UNEP POPs accumulation. HCB was not registered as a pesticide in Japan, but it was found to be about 0.7–1.1% of the derivative of consumed pentachloronitrobenzene, 0.1–9.1% impurities of trichloro terphenyls, and 0.4% in PCP as trace material (Tsuchida et al. 1987; Uemura et al. 2002). HCB also had been found in incinerator exhaust gas (IEG) as well as from IWI. In the IEG, HCB was also found to have a significant linear relationship with PCDDs, with r2 = 0.95. These are the possible explanations for occurrence of HCB in cormorants and fish in this study.
Cyclodienes
The cyclodienes, such as aldrin, dieldrin, and endrin, were also analyzed for the first time in cormorants and fish using HRGC-HRMS. Dieldrin was the predominant contaminant that accumulated in cormorant liver, egg, and fish, respectively (Table 5). The percentage contribution of cyclodienes to the total UNEP POPs was 0.5 to 4.1%, with maximum levels noted in fish and lower contributions found in cormorant eggs. In Japan, 3252, 682, and 1470 tons of aldrin, dieldrin, and endrin, respectively, were used between 1958 and 1972 (EHDJ 2002). Furthermore, sporadic use of imported cyclodienes in Japan contributed to their occurrence in cormorant organs and fish. In most biological tissues, the aldrin could be metabolized into dieldrin (Muralidharan 1993). The other study demonstrated that in biological systems, dieldrin was retained in greater amounts and produced mortality at certain elevated concentrations (Olsen et al. 1992). There are no study reports of liver and egg concentrations of cyclodienes in cormorants from Japan. Dieldrin concentrations in eggs of cormorants from the United States were in the range of 30–1,300 ng/g wet wt (Custer et al. 1997, 1999; Meadows et al. 1996). The wet weight concentrations of dieldrin in cormorant egg in this study are 12–52 ng/g, which are several orders of magnitude lower than cormorant egg from the United States.
Chlordanes
The mean concentrations of chlordanes in cormorant liver, egg, and fish are shown in Table 5. The percentage contribution of chlordanes to the total UNEP POPs was 1.9–22%, with maximum concentrations noted in fish and minimum contributions noted in cormorant eggs. Oxychlordane was the predominant compound followed by cis-nonachlor, heptachlor epoxide, cis-chlordane, trans-nonachlor, trans-chlordane, and heptachlor in cormorant liver and egg. In fish, trans-nonachlor was the predominant compound followed by cis-nonachlor, cis-chlordane, oxychlordane, trans-chlordane, heptachlor epoxide, and heptachlor. A similar chlordane composition was also noted in great cormorants and fish from Lake Biwa and Tokyo Bay in Japan (Guruge et al. 1997). In biological samples, cis/trans-nonachlor could be converted as oxychlordane (Tashiro and Matsumura 1978). In vivo and in vitro studies determined that the immediate major metabolic product of trans-nonachlor is trans-chlordane, which is further converted to 1,2-dichlorochlordene and to oxychlordane. Likewise, in biological samples, heptachlor will be metabolized to heptachlor epoxide during enzymatic degradations. The major metabolic products of heptachlor were heptachlor epoxide, 1-exo-hydroxyheptachlor epoxide, and 1,2-dihydroxydihydrochlordene. The accumulation difference in cormorants and fish could be explained as different metabolic capacity by these animals.
DDTs
Mean concentrations of DDT and its metabolites in cormorant organs and fish are summarized in Table 5. The percent contribution of DDTs was 24 to 39% of the total UNEP POPs, which is greater than any other compounds analyzed. p,p′-DDE had a greater composition of the total DDTs, constituting more than 95% of the total DDT load. The higher composition of p,p′-DDE in most of the animals clearly suggests the greater ability to transform p,p′-DDT into p,p′-DDE, as noted in several wildlife samples (Senthil Kumar et al. 2001a). The relatively larger proportion of p,p′-DDT, which is the major constituent (80%) of the technical DDT mixture in fish, suggests its smaller metabolic capacity. DDT was used in Japan starting in September 27, 1948 and was banned on May 1, 1971 (Uemura et al. 2002). The continued occurrence of DDTs in cormorants even after banning (in the 1970s) in Japan suggests its greater persistence in the environment.
The eggshell thickness and embryonic development is a biomarker for the p,p′-DDE concentrations. Although the shell thicknesses were not measured in this study, the degree of egg shell thinning was assessed from p,p′-DDE concentrations in whole body homogenates (Tanabe et al. 1998; Senthil Kumar et al. 2001a) or in livers using the effective concentrations in birds that were reported in the existing literatures. For example, the average concentrations of p,p′-DDE at 20–1,000 μg/g on a lipid weight basis in the liver of birds was considered to pose a threat to individual bird reproduction and therefore to the population as a whole (Koeman et al. 1973; Platteeuw et al. 1995). The lipid-normalized concentrations of p,p′-DDE in 9 of 10 cormorant livers in this study was greater than 21 μg/g lipid, which is in the range of those values that may cause reproductive abnormalities.
Hazard estimation for eggs can be explained as approximately 5% eggshell thinning or more, reported to occur at a concentration of about 4 μg/g (wet wt) of p,p′-DDE in eggs (Dirksen et al. 1995). Newton (1988) stated that p,p′-DDE concentrations greater than 3 μg/g (wet wt) in peregrine falcons resulted in reduced breeding success. Concentrations of p,p′-DDE in cormorant eggs were 520 to 3700 ng/g (wet wt). If effective concentrations of 3 μg/g (wet wt) taken into consideration, 2 of 10 cormorant eggs analyzed in this study may be close to the threshold of risk. However, species-specific sensitivity for p,p′-DDE was also taken into account for the risk estimations.
Correlations
According to biologists’ information, the cormorant feeds mainly on gizzard shad as its major diet (Watanabe et al. 2004). Consequently, a correlation test was performed for total OCPs in liver and egg versus gizzard shad (Figure 2a,b). Fish–cormorant total OCP concentrations were positively correlated with liver and egg samples (Figure 2a) when one elevated liver sample was included for the correlation test. Removal (considered to outlier) of individual liver sample provides weak correlation (Figure 2b). Therefore, biomagnification of OCPs was not only from gizzard shad analyzed in this study. In addition, great cormorants feed not only on gizzard shad but also on other aquatic biota, and therefore all cormorant feed items should be included for biomagnification discussions.

Correlation of OCPs between cormorant organs and their diet, fish gizzard shad. Graph in (a) shows all samples (including one elevated concentration of liver), whereas graph in (b) shows outlier of elevated liver sample).
Dioxin-like PCBs
The dioxin-like PCBs were second predominant accumulants with 12% (liver) and 38% (egg) contribution to the total UNEP POPs measured in cormorant organs (Table 6). In general, PeCB-118 was the major accumulant followed by PeCB-105, HpCBs-180/170, HxCB-156, and other congeners for mono- and di-ortho PCBs in both liver and egg. Among non-ortho PCBs, PeCB-126 was the predominant contaminant followed by HxCB-169, TeCB-81 and TeCB-77 in either cormorant organ. These congener-specific accumulation patterns were similar to those studies available for cormorants (Williams et al. 1995; Sanderson et al. 1994; van den Berg et al. 1994; Meadows et al. 1996; Guruge et al. 2000). These studies reflect the fact that although technical formulations of PCBs vary between countries (for example, Kanechlor in Japan and Aroclor in United States), the contamination patterns were similar.
Contamination profiles of PCBs were contrary to those of OCPs, which mainly accumulated in liver rather than in egg. Elevated accumulation of PCBs in eggs suggests its efficient transfer from mother to egg. Furthermore, it is worth indicating that eggs were collected at Odaiba of metropolitan Tokyo, which considered to contaminate with PCBs (Senthil Kumar et al. 2002a) because of major industrialization in this area. The liver samples were collected in the Sagami River at Kanagawa Prefecture, where industrial abundance is comparatively less than Tokyo but extensive agricultural activities occur and thus comparison between organs should not be done. Comparison of liver and egg concentrations in this study is not meaningful because of geographical variation in sampling. These are the possible discrepancies for contamination differences between egg and liver.
PCDD/DFs
The PCDD/DFs were the lowest accumulants that contributed 0.01% (liver) and 0.003% (egg) to the total UNEP POPs measured in organs of cormorants (Table 6). Contamination profiles of PCDD/DFs in liver and egg were similar to those of OCPs, which mainly accumulated in liver rather than in egg. 23478-PeCDF was the prevalent congener, followed by 123678-HxCDD, 12378-PeCDD, 234678-HxCDF, OCDD, and so on. The congener-specific pattern providing the PCDD/DF sources were from PCP, CNP, as well as combustion sources (Masunaga et al. 2001a,b; Yao et al. 2002). These studies also stated that herbicide-derived PCDD/DFs remaining in agricultural land in Japan would continue to run off and pollute the aquatic wildlife and environment for prolonged period (Senthil Kumar et al. 2003a).
PCDD/DF concentrations in birds were compared with studies abroad reported to date. The lipid-based concentrations of dioxin-like PCBs and PCDD/DFs in cormorants in this study were greater than those in eggs of southern polar skua and penguin eggs (Senthil Kumar et al. 2002b), liver of predator birds from India (Senthil Kumar et al. 2001b), blood of vultures from the United States (Senthil Kumar et al. 2003b), eggs of common terns from St. Mary’s River, Great Lakes, USA (Senthil Kumar et al. 2003c), and yolk sac of cormorants from The Netherlands (Van den Berg et al. 1994, 1995). However, observed dioxin-like PCBs and PCDD/DFs in this study were slightly higher than in livers of white-tailed sea eagles from Germany and Poland (Kannan et al. 2003) but were similar to those in liver of double-crested cormorants and bald eagles from Michigan, USA (Kannan et al. 2001; Senthil Kumar et al. 2002c), and predator and aquatic birds from Japan (Senthil Kumar et al. 2002a; Iseki et al. 2001a,b; Kang et al. 2002). Based on these observations, double-crested cormorants in the Great Lakes and great cormorants in Japan accumulated elevated PCDD/DF concentrations.
Temporal trend
Temporal trend studies are useful tool in elucidating past history, present status, and predicting the future trend of contamination by persistent, bioaccumulative and toxic chemicals. Temporal trend monitoring data are valuable in determining whether legislative actions taken to reduce the degree of pollution by certain chemicals really have had the intended effect. Temporal trend studies also provide useful information on the effect of a suspected chemical on an animal population. If the levels decrease, but the ecological effect remains, then there is a chance that another contaminant, not originally suspect is solely or partly responsible for it. Considering those facts, we compared our liver OCPs and liver/egg PCB-118 to the suitable literatures (Guruge et al. 1997, 2000) for the years 1993, 1994, and 1998 from Lake Biwa and Tokyo (Figure 3). Temporal trend results show an apparent decrease of HCHs and HCB, in livers of cormorants especially, in Tokyo Shinobazu Pond cormorants collected in 1994. Chlordane (sum of five chlordane compounds except heptachlor and its epoxide) concentrations in cormorants from Japan collected in Shinobazu Pond, Tokyo in 1994 (Guruge et al. 1997), were slightly higher (Figure 3) than those in our study, in which slower degradation of chlordane compounds prevailed in the Japanese environment. Chlordane compounds were used as termite control until 1986 (EHDJ, 2002), which is considered to be a possible explanation of its slower decline rates than other OCPs that were banned in the 1970s. Furthermore, it should be explained that mean concentrations of chlordane compounds were higher in the fish tissue than in cormorant livers. Concentrations of DDTs in Tokyo cormorants collected in 1994 were compared (Figure 3), which is two times greater than those in our study. These results again corroborate degradation of DDTs in the Japanese environment. Because in this study only dioxin-like PCBs were analyzed, the comparison of PCB-118 was possible with Guruge et al. (2000), since this congener was shown to accumulate at more elevated levels than the other PCB congeners in cormorants. The PeCB-118 in liver of cormorant from Tokyo collected in 1994 (Guruge et al. 1997) were greater than the levels noticed in present study (Figure 3). The eggs collected in present study and in Guruge et al. (2000) were from the same years (1998) and thus the temporal trend was not pronounced. Altogether, the temporal trend results in liver with a limited number of samples imply a gradual decrease of POPs in Japan.

Trend analysis of organochlorine pesticides and PCB-118 in cormorants from Japan between 1993 and 2001.
Toxic equivalency
Dioxin-like PCBs and PCDD/DFs TEQ concentrations were calculated based on WHO-TEF proposed for birds (Table 6). The majority of the TEQ was contributed by non-ortho PCBs followed by PCDFs, PCDDs, and mono-ortho PCBs in liver and mono-ortho PCBs, PCDDs, and PCDFs in eggs (Figure 4). Non-ortho PeCB-126 alone contributed more than 35% of the TEQ followed by 23478-PeCDF (>30%), non-ortho TeCB-81 (>5%), 12378-PeCDD (>5%), 234678-HxCDF (>5%), mono-ortho PCBs such as PeCB-105, HxCB-156, PeCB-118 in liver and egg, with the collective contribution by the aforementioned congeners ranging from 82% to 92%. Concentrations of PeCB-126 were greater than TeCB-77 in liver and egg. This pattern is different from what is observed for technical PCB mixtures, which are the major sources of exposure of non-ortho PCBs in wildlife (Senthil Kumar et al. 1999a,b). Higher concentrations of PeCB-126 than TeCB-77 in liver suggest metabolism of PCB-77 in birds (Senthil Kumar et al. 2002c). The higher PCB load also increases the proportion of IUPAC 169/126 ratio and also encourages strong enzyme induction (Kannan et al. 1993). The strong induction of drug-metabolizing enzymes results in higher biotransformation of TeCB-77 to PeCB-126 rather than HxCB-169, hence the CB 169/126 ratio could be increased (Senthil Kumar et al. 1999a,b, 2002c). This observation may suggest an alteration in cytochrome P450 enzyme–related metabolism of several endobiotics, which could disrupt the endocrine system.

TEQ contribution by PCDDs, PCDFs, and non- and mono-ortho PCBs in livers and eggs of great cormorants.
Toxic threshold
The toxic threshold for avian species has been reported elsewhere. The no-observed-effect-level (NOAEL) of 100 pg/g TEQs and low-observed-effect-level (LOAEL) of 210 pg/g TEQs on a wet wt basis are suggested for bald eagle eggs (Elliott and Norstrom, 1998). The proposed toxic threshold values were much greater than the wet wt TEQ observed in great cormorant egg with mean (ranges) of 4.5 (1.6–9.3) pg/g in this study. A lowest-observed effect level (LOEL) of 25 ng TEQ/g in liver on a lipid basis has been suggested for CYP1A induction and 50% reduction of plasma thyroxin levels in common tern (Sterna hirundo) chicks (Bosveld et al., 2000). The mean liver TEQ concentrations were 17 with the ranges of 4–31 ng/g lipid with 3 birds containing more than 25 and thus CYP1A or ethoxyresorufin-O-deethylase (EROD) was expected in these birds; however, we do not know the age of the birds analyzed in this study and thus CYP1A interpretation should be considered with caution.
The induction of isoenzyme CYP1A1, which is a highly sensitive biomarker for dioxin-like compounds, has been measured by measuring EROD induction in birds. Few investigations suggested that EROD activity was not induced to levels greater than the background in the dose group receiving only 0.1 ng PeCB-126/g, but the group receiving a dose of 1 ng PeCB-126/g was induced 2.4-fold relative to background concentrations in the laboratory (Bosveld et al. 2000). The PeCB-126 concentrations in liver and egg of cormorants analyzed in this study were several-fold greater than that which induced EROD activity.
Conclusion
In summary, this is the first report attempting to establish 11 among 12 UNEP POPs in HRGC-HRMS. Based on the results, ultratrace (femtograms) of UNEP POPs can be determined using HRGC-HRMS with inclusion of isotope-labeled standards. Except PCDD/DFs, other UNEP POPs are still reported to occur at parts per billion levels. DDTs are predominant contaminants, followed by dioxin-like PCBs, chlordanes, cyclodienes, HCB, and HCHs, whereas minimum accumulation was found for PCDD/DFs. Continued occurrence of a considerable amount of some OCPs and elevated exposures of PCBs in Japan are of major concern. Nevertheless, temporal trends of OCPs and PeCB-118 in limited samples showed a decreasing trend between 1994 and 2000. Current concentrations of DDTs, dioxin-like PCBs, PCDD/DFs, and their TEQ in liver and egg were still close to posing a substantial impact on health in birds.
References
JI Baker RA Hites (2000) ArticleTitleIn combustion the major source of polychlorinated dibenzo-p-dioxins and dibenzofurans to the environment? A mass balance investigation Environ Sci Technol 34 2879–2886 Occurrence Handle1:CAS:528:DC%2BD3cXjvVait7g%3D
LS Birnbaum J Tuomisto (2000) ArticleTitleNon-carcinogenic effects of TCDD in animals Food Addit Contam 17 275–288 Occurrence Handle1:CAS:528:DC%2BD3cXktl2qtbc%3D Occurrence Handle10912242
ATC Bosveld R Nieboer J Bont Particlede AJ Murk LA Feyk JP Giesy M Berg ParticleVan den (2000) ArticleTitleBiochemical and developmental effects of dietary exposure to polychlorinated biphenyls 126 and 153 in common tern chicks (Sterna hirundo) Environ Toxicol Chem 19 719–730 Occurrence Handle1:CAS:528:DC%2BD3cXhtlymsr0%3D
TJ Boudewijn S Dirksen (1995) ArticleTitleImpact of contaminants on the breeding success of the cormorant Phalacrocorax carbo sinensis in The Netherlands Ardea 83 325–338
TW Custer CM Custer KL Stromborg (1997) ArticleTitleDistribution of organochlorine contaminants in double-crested cormorant eggs and sibling embryos Environ Toxicol Chem 16 1646–1649 Occurrence Handle1:CAS:528:DyaK2sXlt1Sjt74%3D
TW Custer CM Custer RK Hines S Gutreuter KL Stromborg PD Allen MJ Melancon (1999) ArticleTitleOrganochlorine contaminants and reproductive success of double-crested cormorants from Green Bay, Wisconsin, USA Environ Toxicol Chem 18 1209–1217 Occurrence Handle1:CAS:528:DyaK1MXjt1SrtL8%3D
TW Custer CM Custer RK Hines S Stromborg PD Allen MJ Melancon DS Henshel (2001) ArticleTitleOrganochlorine contaminants and biomarker response in double-crested cormorants nesting in Green Bay, and Lake Michigan, Wisconsin, USA Arch Environ Contam Toxicol 40 89–100 Occurrence Handle1:CAS:528:DC%2BD3MXhs1CgsLs%3D Occurrence Handle11116344
S Dirksen TJ Boudewijn LK Slager RG Mes MJM Schaick Particlevan P Voogt Particlede (1995) ArticleTitleReduced breeding success of cormorants (Phalacrocorax carbo sinensis) in relation to persistent organochlorine pollution of aquatic habits in the Netherlands Environ Pollut 88 119–132 Occurrence Handle1:CAS:528:DyaK2MXjvFKlsrw%3D Occurrence Handle15091551
EHDJ (2002) The environmental monitoring report on the persistent organic pollutants (POPs) in Japan. Environmental Health Department of Japan, Ministry of the Environment, Government of Japan http://www.env.go.jp/en/
JE Elliott RJ Norstrom (1998) ArticleTitleChlorinated hydrocarbon contaminants and productivity of bald eagle populations on the Pacific Coast of Canada Environ Toxicol Chem 17 1142–1153 Occurrence Handle1:CAS:528:DyaK1cXjsV2qsb4%3D
M Feeley A Brouwer (2000) ArticleTitleHealth risks to infants from exposure to PCBs, PCDDs and PCDFs Food Addit Contam 17 325–333 Occurrence Handle1:CAS:528:DC%2BD3cXktl2qur0%3D Occurrence Handle10912246
KS Guruge S Tanabe M Fukuda S Yamagishi R Tatsukawa (1997) ArticleTitleAccumulation pattern of persistent organochlorine residues in Common cormorants (Phalacrocorax carbo) from Japan Mar Poll Bull 34 186–193 Occurrence Handle1:CAS:528:DyaK2sXjt1aks7s%3D
KS Guruge S Tanabe M Fukuda (2000) ArticleTitleToxic assessment of PCBs by the 2,3,7,8-tetrachlorodibenzo-p-dioxin equivalent in common cormorant (Phalacrocorax carbo) from Japan Arch Environ Contam Toxicol 38 509–521 Occurrence Handle1:CAS:528:DC%2BD3cXivVKgsbs%3D Occurrence Handle10787103
M Hosomi T Matsuo S Dobashi S Katou H Abe (2003) ArticleTitleSurvey of dioxins in Tokyo Bay bottom sediment Mar Pollut Bull 47 68–73 Occurrence Handle1:CAS:528:DC%2BD3sXkt1Gitrg%3D Occurrence Handle12787599
F Iino T Imagawa BK Gullett (2000) ArticleTitleDechlorinated-controlled polychlorinated dibenzofuran isomer patterns from municipal waste incinerators Environ Sci Technol 34 3143–3147 Occurrence Handle1:CAS:528:DC%2BD3cXkt1yrtr4%3D
N Iseki S Hayama S Masunaga J Nakanishi (2001a) ArticleTitleConcentrations of PCDDs, PCDFs and dioxin-like PCBs; their biomagnification factor in common cormorants (Phalacrocorax carbo) from Japan Organohalogen Compounds 51 332–335 Occurrence Handle1:CAS:528:DC%2BD38Xnt1yju7g%3D
N Iseki T Iizuka S Hayama S Masunaga J Nakanishi (2001b) ArticleTitleAccumulation profiles and burdens of dioxins, furans and dioxin-like PCBs in common cormorants (Phalacrocorax carbo) from Japan Organohalogen Compounds 51 336–339 Occurrence Handle1:CAS:528:DC%2BD38Xnt1yju7k%3D
YS Kang M Yamamuro S Masunaga J Nakanishi (2002) ArticleTitleSpecific biomagnification of polychlorinated dibenzo-p-dioxins and dibenzofurans in tufted ducks (Athya fuligula), common cormorants (Phalacrocorax carbo) and their prey from Lake Shinji, Japan Chemosphere 46 1373–1382 Occurrence Handle1:CAS:528:DC%2BD38XhvVChsr8%3D Occurrence Handle12002464
K Kannan S Tanabe A Borrell A Aguilar S Focardi R Tatsukawa (1993) ArticleTitleIsomer-specific analysis and toxic evaluation of polychlorinated biphenyls in striped dolphins affected by an epizootic in the western Mediterranean Sea Arch Environ Contam Toxicol 25 227–233 Occurrence Handle1:CAS:528:DyaK3sXlsVKju7Y%3D Occurrence Handle8368865
K Kannan K Hilscherova T Imagawa N Yamashita LL Williams JP Giesy (2001) ArticleTitlePolychlorinated- naphthalenes, -biphenyls, -dibenzo-p-dioxins and –dibenzofurans in double-crested cormorants and herring gulls from Michigan waters of Great Lakes Environ Sci Technol 35 441–447 Occurrence Handle1:CAS:528:DC%2BD3cXovFSjs7g%3D Occurrence Handle11351712
K Kannan K Senthil Kumar H Nakata J Falandysz G Oehme S Masunaga (2003) ArticleTitlePolychlorinated biphenyls, dibenzo-p-dioxins, dibenzofurans and p,p’-DDE in livers of white-tailed sea eagles from eastern Germany, 1979–1998 Environ Sci Technol 37 1249–1255 Occurrence Handle1:CAS:528:DC%2BD3sXhsVaqurg%3D
JH Koeman HCW Vanvelzen-blad R Devries JG Vos (1973) ArticleTitleEffects of PCB and DDE in cormorants and evaluation of PCB residues from an experimental study J Reprod Fertil Suppl 19 353–364 Occurrence Handle1:CAS:528:DyaE2cXksVKitrc%3D Occurrence Handle4206409
JM Larson WH Karasov L Sileo KL Stromborg BA Hanbidge JP Giesy PD Jones DE Tillitt DA Verbrugge (1996) ArticleTitleReproductive success, developmental anomalies, and environmental contaminants in double-crested cormorants (Phalacrocorax auritus) Environ Toxicol Chem 15 553–559 Occurrence Handle1:CAS:528:DyaK28XitVClsb8%3D
S Masunaga T Takasuga J Nakanishi (2001a) ArticleTitleDioxin and dioxin-like PCBs impurities in some Japanese agrochemical formulations Chemosphere 44 873–885 Occurrence Handle1:CAS:528:DC%2BD3MXkvVehtbg%3D
S Masunaga Y Yao I Ogura S Nakai Y Kanai M Yamamuro J Nakanishi (2001b) ArticleTitleIdentifying sources and mass balance of dioxin pollution in Lake Shinji Basin, Japan Environ Sci Technol 35 1967–1973 Occurrence Handle1:CAS:528:DC%2BD3MXisVansLs%3D
H Matsukami K Senthil Kumar M Yamashita E Ohi T Takasuga (2004) ArticleTitleAnalytical methodology development for toxaphene by high resolution gas chromatography/high resolution mass spectrometry with electron capture negative ionization Organohalogen Compounds 66 219–225
JC Meadows DE Tillitt TR Schwartz DJ Schroeder KR Echols RW Gale DC Powell SJ Bursian (1996) ArticleTitleOrganochlorine contaminants in double-crested cormorants from Green Bay, WI: I. Large-scale extraction and isolation from eggs using semi-permeable membrane dialysis Arch Environ Contam Toxicol 31 218–224 Occurrence Handle1:CAS:528:DyaK28XlsVemsLw%3D Occurrence Handle8781072
S Muralidharan (1993) ArticleTitleAldrin poisoning of sarus cranes (Grus antigone) and a few granivorous birds in Keoladeo National Park, Bharatpur, India Ecotoxicol 2 196–202 Occurrence Handle1:CAS:528:DyaK2cXht1OgsrY%3D
I Newton (1988) ArticleTitleDetermination of critical pollutant levels in wild populations, with examples from organochlorine insecticides in birds of prey Environ Pollut 55 29–40 Occurrence Handle1:CAS:528:DyaL1MXht1yisg%3D%3D Occurrence Handle15092513
P Olsen B Emison N Mooney N Brothers (1992) ArticleTitleDDT and dieldrin: effects on resident peregrine falcon populations in south-eastern Australia Ecotoxicol 1 89–100 Occurrence Handle1:CAS:528:DyaK3sXhsVWmsrw%3D
M Platteeuw MR Eerden Particlevan K av de Guchte (1995) ArticleTitleVariation in contaminant content of livers of cormorants Phalacrocorax carbo sinensis living near polluted sedimentation area in Lake Ijsselmeer in the Netherlands Ardea 83 315–324
DC Powell RJ Aulerich JC Meadows DE Tillitt KL Stromborg TJ Kubiak JP Giesy SJ Bursian (1997) ArticleTitleOrganochlorine contaminants in double-crested cormorants from Green Bay, Wisconsin: II. Effects of an extract derived from cormorant eggs on the chicken embryo Arch Environ Contam Toxicol 32 316–322 Occurrence Handle1:CAS:528:DyaK2sXislWlu7g%3D Occurrence Handle9096082
JT Sanderson RJ Norstrom JE Elliott LE Hart KM Cheng GD Bellward (1994) ArticleTitleBiological effects of polychlorinated dibenzo-p-dioxins, dibenzofurans, and biphenyls in double-crested cormorant chicks (Phalacrocorax auritus) J Toxicol Environ Health 41 247–265 Occurrence Handle1:CAS:528:DyaK2cXhvVCqt78%3D Occurrence Handle8301702
K Senthil Kumar M Watanabe K Kannan AN Subramanian S Tanabe (1999a) ArticleTitleIsomer-specific patterns and toxic assessment of polychlorinated biphenyls in resident, wintering migrant birds and bat collected from South India Toxicol Environ Chem 71 221–239 Occurrence Handle1:CAS:528:DyaK1MXnsVWntbY%3D
K Senthil Kumar K Kannan RK Sinha S Tanabe JP Giesy (1999b) ArticleTitleBioaccumulation profiles of polychlorinated biphenyl congeners and organochlorine pesticides in Ganges River dolphins Environ Toxicol Chem 18 1511–1520 Occurrence Handle1:CAS:528:DyaK1MXktVequ7c%3D
K Senthil Kumar K Kannan S Tanabe AN Subramanian (2001a) ArticleTitleAccumulation of persistent organochlorine pesticides and PCBs in sediments, aquatic biota, birds, eggs and bat collected from South India Environ Sci Pollut Res 8 35–47 Occurrence Handle1:CAS:528:DC%2BD3MXit1eksrg%3D
K Senthil Kumar K Kannan ON Paramasivan VP Shanmugasundaram J Nakanishi S Masunaga (2001b) ArticleTitlePolychlorinated dibenzo-p-dioxins, dibenzofurans, and polychlorinated biphenyls in human tissues, meat, fish and wildlife samples from India Environ Sci Technol 35 3448–3455 Occurrence Handle10.1021/es010555+
K Senthil Kumar N Iseki SI Hayama J Nakanishi S Masunaga (2002a) ArticleTitlePolychlorinated dibenzo-p-dioxins, dibenzofurans and dioxin-like polychlorinated biphenyls in livers of birds from Japan Arch Environ Contam Toxicol 42 244–255 Occurrence Handle1:STN:280:DC%2BD38%2FoslWhsg%3D%3D
K Senthil Kumar K Kannan S Corlsolini T Evans JP Giesy J Nakanishi S Masunaga (2002b) ArticleTitlePolychlorinated dibenzo-p-dioxins, dibenzofurans and polychlorinated biphenyls in polar bear, penguin and polar skua Environ Pollut 119 151–161 Occurrence Handle10.1016/S0269-7491(01)00332-3
K Senthil Kumar K Kannan JP Giesy S Masunaga (2002c) ArticleTitleDistribution and elimination of polychlorinated dibenzo-p-dioxins, dibenzofurans, biphenyls and p,p’-DDE in tissues of bald eagles from the upper peninsula of Michigan Environ Sci Technol 36 2789–2796 Occurrence Handle10.1021/es0114660
K Senthil Kumar N Iseki InstitutionalAuthorNameSunardi S Kasuga T Takasuga S Masunaga (2003a) ArticleTitlePolychlorinated -dibenzo-p-dioxjns, -dibenzofurans and -dioxin-like PCBs in aquatic wildlife of Kasumigaura Lake, Japan Toxicol Environ Chem 85 121–132 Occurrence Handle1:CAS:528:DC%2BD2cXjsVaitr8%3D
K Senthil Kumar WW Bowerman LDV Travis OE Rhodes SuffixJr IL Brisbin SuffixJr T Takasuga S Masunaga (2003b) ArticleTitleChlorinated hydrocarbon contaminants in blood of black and turkey vultures from Savannah River Site of South Carolina, USA Chemosphere 53 173–182 Occurrence Handle1:CAS:528:DC%2BD3sXlvVCmurc%3D
K Senthil Kumar WW Bowerman K Millenbah DA Best T Takasuga S Masunaga (2003c) ArticleTitlePolychlorinated dibenzo-p-dioxins/furans and dioxin-like polychlorinated biphenyls in the eggs of common terns from Lime Island of St. Mary’s River in Michigan, USA Toxicol Environ Chem 85 221–232 Occurrence Handle1:CAS:528:DC%2BD2cXjt1ymtLw%3D
MH Sweeney InstitutionalAuthorNameMocarelli (2000) ArticleTitleHuman health effects after exposure to 2,3,7,8-TCDD Food Addit Contam 17 303–316 Occurrence Handle1:CAS:528:DC%2BD3cXktl2qur8%3D Occurrence Handle10912244
T Takasuga M Yasuda F Yamada K Senthil Kumar (2003) ArticleTitleBioaccumulation profiles of UNEP’s POPs in terrestrial wildlife in some parts of Japan Organohalogen Compounds 64 360–365 Occurrence Handle1:CAS:528:DC%2BD2cXhs1Omtw%3D%3D
S Tanabe K Senthil Kumar K Kannan AN Subramanian (1998) ArticleTitleAccumulation features of polychlorinated biphenyls and organochlorine pesticides in resident and migratory birds collected from South India Arch Environ Contam Toxicol 34 387– 397 Occurrence Handle1:CAS:528:DyaK1cXislWitro%3D Occurrence Handle9543510
S Tashiro F Matsumura (1978) ArticleTitleMetabolism of trans-nonachlor and related chlordane compounds in rat and man Arch Environ Contam Toxicol 7 113–127 Occurrence Handle1:CAS:528:DyaE1cXltVWhtbY%3D Occurrence Handle666363
DE Tillitt GT Ankley JP Giesy JP Ludwig H Kurita-matsuba DV Weseloh PS Ross A Bishop L Soleo KL Stromborg J Larson TJ Kubiak (1992) ArticleTitlePolychlorinated biphenyl residues and egg mortality in double-crested cormorants from the Great Lakes Environ Toxicol Chem 11 1281–1288 Occurrence Handle1:CAS:528:DyaK38XmtVKlsrw%3D
H Tsuchida Y Hanai F Sagawa T Katou (1987) ArticleTitleThe environmental movement of a nematicide PCNB (quintozene) Bull Instit Environ Sci Technol YNU 14 1–13 Occurrence Handle1:CAS:528:DyaL1MXpsVSitg%3D%3D
Uemura S, Kawamura H, Tsuji M, Tomita S, Maeda S (2002) Dictionary of pesticides toxicity, Tokyo, Japan 183–186in Japanese
UNEP Chemicals GMN (2001) UNEP Governing Council Decision 19/13C (1997) invited UNEP to convene an Intergovernmental Negotiating Committee (INC) to prepare an international legally binding instrument on persistent organic pollutants (POPs). Stockholm Convention Meeting conducted by United Nations Environment Programme [UNEP] in May 2001 (http://www.pops.int/)
UNEP Chemicals GMN (2002) First Meeting of the Advisory Group to the Global Network for the Monitoring of Chemicals in the Environment, UNEP Chemicals, Geneva, Switzerland, 13 to 14 May 2002. (http://www.chem.unep.ch/gmn/Files/GMN_AdvGrp02_report_final.pdf)
M Berg ParticleVan den BLHJ Craane T Sinnige SV Mourik S Dirksen T Boudewijn M Gaag Particlevan der IJ Lutke-Schipholt B Spenkelink A Brouwer (1994) ArticleTitleBiochemical and toxic effects of polychlorinated biphenyls (PCBs), dibenzo-p-dioxins (PCDDs) and dibenzofurans (PCDFs) in the cormorant (Phalacrocorax carbo) after in ovo exposure Environ Toxicol Chem 13 803–816
M Berg ParticleVan den BLHJ Craane SV Mourik A Brouwer (1995) ArticleTitleThe (possible) impact of chlorinated dioxins (PCDDs), dibenzofurans (PCDFs) and biphenyls (PCBs) on the reproduction of the cormorant Phalacrocorax carbo-An ecotoxicological approach Ardea 83 299–313
M Berg ParticleVan den A Birgelen ParticleVan L Birnbaum B Brouwer G Carrier Y Dragan W Farland M Feeley P Furst CL Galli J Greig Y Hayashi M Kogevinas Y Kurokawa JC Larsen AKD Liem F Matsumura P Mocarelli MR Moore RC Newhook RE Peterson L Poellinger C Portier WJ Rogan D Schrenk MH Sweeney C Tohyama J Tuomisto J Waters M Zeilmaker (2000) ArticleTitleConsultation on assessment of the health risk of dioxins; re-evaluation of the tolerable daily intake (TDI): executive summary Food Addit Contam 17 223–240 Occurrence Handle10.1080/026520300283315
K Watanabe K Senthil Kumar S Masunaga T Takasuga N Iseki M Morita (2004) ArticleTitleBrominated organic compounds in the liver and egg of the common cormorants (Phalacrocorax carbo) from Japan Environ Sci Technol 38 4071–4077 Occurrence Handle10.1021/es0307221 Occurrence Handle1:CAS:528:DC%2BD2cXltFWht70%3D Occurrence Handle15352443
LL Williams JP Giesy DA Verbrugge S Jurzysta K Stromborg (1995) ArticleTitlePolychlorinated biphenyls and 2,3,7,8-tetrachlorodibenzo-p-dioxin equivalents in eggs of double-crested cormorants from a colony near Green Bay, Wisconsin, USA Arch Environ Contam Toxicol 29 327–333 Occurrence Handle10.1007/BF00212497 Occurrence Handle1:CAS:528:DyaK2MXnvVGqsL0%3D Occurrence Handle7487156
Yamashita M, Takemori H, Senthil Kumar K, Takasuga (2002) Analysis of Stockholm Convention priority POPs with high sensitivity and high quality using 13C-isotope standards and HRGC-HRMS: clean-up and analysis. The 5th Annual Meeting of Japan Society of Endocrine Disruptors Research 25–26th November 2002, p 157
Y Yao S Masunaga H Takada J Nakanishi (2002) ArticleTitleIdentification of polychlorinated dibenzo-p-dioxins, dibenzofuran, and coplanar polychlorinated biphenyl sources in Tokyo Bay, Japan Environ Toxicol Chem 21 991–998 Occurrence Handle10.1897/1551-5028(2002)021<0991:IOPDPD>2.0.CO;2 Occurrence Handle1:CAS:528:DC%2BD38XktlWltbw%3D Occurrence Handle12013146
Yasuda M, Yamada F, Kawaji N, Yamazaki K, Nakajima K, Ishizuka T, Takasuga T Senthil Kumar K (2003) Characteristics of POPs in wildlife in Kanto area Japan. 12th Annual Meeting of the Japan Society for Environmental Chemistry, 24–28 June 2003, Niigata, Japan
Acknowledgments
We would like to thank Mr. Etsumasa Ohi and Miss Michiko Yamashita, Shimadzu Techno Research Inc., Kyoto, Japan for the help during analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kumar, K.S., Watanabe, K., Takemori, H. et al. Analysis of UNEP Priority POPs Using HRGC-HRMS and Their Contamination Profiles in Livers and Eggs of Great Cormorants (Phalacrocorax carbo) from Japan. Arch Environ Contam Toxicol 48, 538–551 (2005). https://doi.org/10.1007/s00244-004-0030-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00244-004-0030-3