
Abstract
The effects of different concentrations of selenite (2–30 μM) and selenate (2–60 μM) on biomass production, leaf area, and concentrations of photosynthetic pigments in lettuce plants were investigated. On the basis of the obtained results, the threshold of toxicity for the selenite and selenate has been designated. The toxicity thresholds for selenite and selenate were determined at concentrations of 15 and 20 μM, respectively. Next, four selenium (Se) concentrations (2, 4, 6 or 15 μM), below or near the toxicity boundary, have been selected for the lettuce biofortification experiment. In the biofortified plants, the oxidant status (levels of lipid peroxidation and H2O2 concentrations), as well as Se and sulphur (S) accumulation were analysed. In the edible parts of the lettuce, the Se concentration was higher for selenate presence compared to selenite; however, this difference was not as obvious as it was noted in the case of the roots, where selenite application caused the high accumulation of Se. An application of 15 μM Se as selenite caused a decline in the biomass and an intensification of prooxidative processes in the plant’s tissues and as toxic should be excluded from further biofortification experiments. These results indicate that an application of either selenate or selenite to the nutrient solution at concentrations below 15 μM can be used for biofortification of lettuce with Se, evoking better plant growth and not inducing significant changes in the oxidant status, the concentration of assimilation pigments and S accumulation.
Similar content being viewed by others

Introduction
Humans require more than 22 mineral elements for proper health and development, which can be provided by an appropriate diet. Nevertheless, it is estimated that over 60 % of the world’s population are iron (Fe) deficient, over 30 % are zinc (Zn) deficient, 30 % are iodine (I) deficient and 15 % are selenium (Se) deficient. This situation is attributed mainly to crop production in areas with naturally low mineral concentration or phytoavailability and/or consumption of crops with inherently low tissue mineral concentrations, compounded by a low consumption of fish or animal products. Mineral malnutrition can be reduced through well-chosen dietary diversification, mineral supplementation, food fortification and/or by increasing the bio-available mineral concentration in edible crops (biofortification) (White and Broadley 2009).
Se is a biologically-active element that can form direct Se–C bonds, occurring in a range of essential organic compounds for humans and animals, including selenoacids and selenoproteins (Broadley et al. 2006). Because either insufficient or excessive Se intake can have dramatic consequences for health, this element is often described as a ‘double-edged sword’. Se is incorporated into the food chain mainly via crops and therefore the Se status of the food chain is dependent on Se level in the soil. In areas where soils are naturally low in bio-available Se, potential Se deficiencies may cause serious health problems (Hartikainen 2005). The Se level in the soils of many European countries generally is low (Rayman 2000, Oldfield 2002). Moreover, the significant impact on reducing Se content in European food was the substitution of North American wheat imports from produce grown within Europe (Bryszewska et al. 2007). The studies performed in Poland, based on Se concentrations in cow’s milk and partly on Se concentrations in grass and in livers of hares, showed that about 75 % of Poland is Se-deficient and only 25 % is adequate, when considering the nutritional requirements of humans and animals (Zachara and Pilecki 2000, Dębski et al. 2001). Because Se deficiency in the diet is a common phenomenon in many countries around the world, plants biofortified with Se are an excellent source of dietary Se than can help to alleviate this problem (Garcia-Banuelos et al. 2011). However, if Se biofortification of crops through fertilisers (agronomic biofortification) is to occur, whether locally or nationally, it issensible to do it as efficiently as possible, especially as Se can be considered as a non-renewable resource which is difficult to recycle (Haug et al. 2007).
Se can exist in the soil solution mainly as selenate (Se VI), selenite (Se IV) and organic forms (SeCys and SeMet), although elemental Se and selenide-Se can also occur depending upon the redox potential of the soil (Terry et al. 2000). Plant roots can uptake this element as selenate, selenite or organo-Se compounds, but cannot take up colloidal elemental Se or metal selenides. Selenate is transported across the plasma membrane by high-affinity sulphate transporters, whilst selenite is thought to be transported by phosphate transporters (White and Broadley 2009) or enters the plant cells passively (Terry et al. 2000). Moreover, recent investigations show that at low pH selenite can enter root cells through an aquaporin channel (Broadley et al. 2012). Although all plants are able to uptake and metabolise Se, however, the question of Se’s essentiality for vascular (higher) plants remains unresolved after nearly 70 years of investigation (Lyons et al. 2009).
Lettuce (Lactuca sativa L.) is the leafy vegetable crop more produced and consumed in the world (Li et al. 2010), and is one of the most commonly grown hydroponic vegetables. This crop can be preferred in Se biofortification programs, as an efficient method of increasing intake of this element by the people in countries where Se level in food is insufficient (Ramos et al. 2010). In regard of the numerous alterations of Se compounds in the soil’s environment (Terry et al. 2000), the stability, mobility and bioavailability of selenates and selenites under an ever changing soil environment is sometimes difficult to predict. Thus, biofortification of hydroponic grown lettuce with Se is a promising and easy technique for increasing Se concentration in the human diet. So in the present study, the effect of the two mineral forms of Se on biofortification efficiency, growth and some physiological parameters of lettuce plants grown in hydroponic cultures has been examined.
Materials and methods
Plant materials, growing conditions, and experimental design
Seeds of butterhead lettuce (L. sativa L. var. capitata) cv. Justyna were sown onto wet quartz sand and germinated at 25 °C for 14–20 days. Then, the seedlings were transferred to 1 dm3 glass jars (two plants each) containing 1.5-fold concentrated Hoagland’s II nutrient solution, supplemented by 2 % ferric citrate and micronutrients in the form of 1 % A–Z solution (Hoagland and Arnon 1950). The pH of the medium was adjusted to 6.0. The nutrient solution was differentiated in regard to the form and concentration of Se: 0, 2, 4, 6, 15, 20, 30, 40 or 60 μM Se applied as selenate (Na2SeO4) or 0, 2, 4, 6, 10, 15, 20, 25 or 30 μM applied as selenite (Na2SeO3). These concentrations were selected based on previous studies and taking into consideration the differentiated toxicity of both Se forms for diverse plant species. Plants were cultured for 14 days in a vegetation chamber (Sanyo MRL 350 HT) under the following conditions: photosynthetic photo flux density of 270 μmol m−2 s−1, 12 h day length, temperature of 22/18 °C (day/night) and relative humidity of 65–70 %. Then, the plant samples were tested for photosynthetic pigments concentration, leaf areas, and biomass accumulation. On the basis of the obtained results, the threshold of toxicity for the studied Se compounds was determined. The toxicity threshold was defined as the lowest concentration of Se that caused a significant decrease in shoots or roots FW compared to the control plants. In the next part of the experiment, the four Se concentrations below or close to the toxicity threshold values (2, 4, 6, 15 μM) were selected for the biofortification experiment. The plants were cultivated in the way described above. After 14 days of plant vegetation under differentiated Se forms and concentrations, the plants were harvested and the levels of lipid peroxidation and H2O2 concentration was evaluated in the leaves and roots. Then, the plant material was dried at 80 °C until a constant mass of the samples was achieved and total Se and S were determined in the shoots and roots.
Analysis of growth parameters and photosynthetic pigments concentrations
The shoots and roots of plants from each jar were harvested separately after 14 days from Se addition. Some growth parameters such as the FW of shoots and roots and leaf areas were measured from the collected plants. Fresh second leaves were scanned using CI-202 laser areametre (CID Bio-Science, USA) and the leaf area was expressed in square centimetres (cm2). Chlorophyll a and b together with all carotenoids (xanthophylls + carotenes) were assayed according to the method of Lichtenthaler and Wellburn (1983). The samples were collected from second leaves and the pigments were extracted from fresh leaves discs via homogenisation in 80 % (v/v) acetone. The absorbance of the resulting extracts was measured at 646, 663, and 470 nm.
Determinations of lipid peroxidation and H2O2 concentration
The thiobarbituric acid (TBA) test is an easy and quick assay for the assessment of lipid peroxidation of cell membranes in which malondialdehyde (MDA) is a by-product of lipid peroxidation in tissue extracts. MDA concentrations were assayed following the method of Heath and Packer (1968) with minor modifications. In order to determine the MDA concentrations, the tissue samples (500 mg) were ground in 4.5 cm3 of 0.1 % (w/v) trichloroacetic acid (TCA) and centrifuged at 10,000 rpm for 10 min. Then, 4 cm3 of 20 % TCA containing 0.5 % of TBA (w/v) was added to 1 cm3 of the obtained supernatant. The mixture of TCA + TBA was enriched using butylated hydroxytoulene (BHT) to prevent MDA formation during the assay. The solutions were heated to 95 °C for 30 min, quickly cooled on ice and re-centrifuged for 10 min. The absorbance of solutions was measured at 532 and 600 nm. The amount of MDA–TBA complex was calculated from the extinction coefficient of 155 mM−1 cm−1.
The level of H2O2 was measured colorimetrically by the modified method of Jana and Choudhuri (1982). H2O2 was extracted by homogenising 250 mg of plant’s tissue with 3 cm3 of phosphate buffer (50 mM, pH 6.5). The homogenate was centrifuged at 6,000 rpm for 25 min. To determine H2O2 concentrations, 3 cm3 of the obtained supernatant was mixed with 1 cm3 of 0.1 % titanium dioxide in 20 % (w/v) H2SO4. Then, the mixture was centrifuged at 6,000 rpm for 15 min. The intensity of the yellow colour of the supernatant was measured at 410 nm. H2O2 level was calculated using the extinction coefficient of 0.28 μM−1 cm−1.
Total Se and S determinations
For the measurement of the total Se concentration, the dried shoot and root samples were subjected to the classic nitric–perchloric mineralisation (HNO3–HClO4; 4:1; v/v) at 210 °C, after which hydride generation atomic absorption spectroscopy (HGAAS) was used to measure the total Se concentrations in the acid digests. This method is based on Se’s conversion to a volatile hydride (SeH2) by the NaBH4 and aspiration into an atomic absorption spectrometer (Perkin Elmer 1100B) fitted with a hydride generation system (Perkin Elmer MHS-10). In brief, after cooling the samples, acid digests were quantitatively transferred to the volumetric flasks, 5 M HCl and deionised water were added, and then heated 30 min at 80 °C to reduce all Se to four oxidation state. After the reduction, all Se was converted to SeH2 with 3 % NaBH4 in 1 % NaOH. The signal was recorded at a wavelength of 196 nm. A certified reference material (WEPAL IPE-157; beech leaf) was used for method validation.
The concentration of total S was estimated using nephelometric method. S mineralisation (the transformation of S-organic to S-sulphate) was performed using NaHCO3. The ground plant material was placed into a crucible, assailed with NaHCO3, and mineralised at 500 °C for 5 h. To the cooled material 25 % HNO3 was added and the dissolved precipitate was quantitatively transferred to the volumetric flasks, which were replenished with deionised water and mixed thoroughly. Next, the appropriate volume of the obtained solution was transferred to a next flask, and 25 % HNO3, 50 % CH3COOH and 85 % H3PO4 were added. The flasks were thoroughly mixed and then BaCl2 in crystals and arabic gum were added. Transmission measurement was done at 490 nm. The standard curve was used to determine the total S concentration of each sample.
All of Se and S analyses were performed in accredited laboratory of Regional Chemical-Agricultural Station in Lublin.
Statistical analysis
The experiment was established in a randomised block design. The values represent the means from three independent repetitions over time. All the obtained data were subjected to a two-way analysis of variance (ANOVA) at 95 % confidence and the significance of differences between mean values were separated using Tukey’s multiple range test.
Results
Threshold of Se toxicity
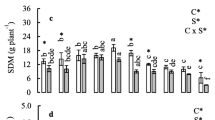
Threshold of Se toxicity, in relation to the chemical form of Se, has been designated on the basis of fresh biomass of plant’s organs and defined as the lowest concentration of Se causing a significant decrease in shoots or roots FW compared to the control value. The biomass of shoots decreased significantly if the selenate and selenite concentration in nutrient solution reached 20 and 15 μM, respectively (Fig. 1a, b). Increasing Se concentrations caused a progressive reduction in FW, both roots and shoots, as well as a significant decline of leaf area. However, at low concentrations Se exerted beneficial effects on the plant’s growth. A significant increase in root and/or shoot FW in the plants supplemented with 6 μM of selenate and 2–10 μM of selenite was identified (Fig. 1a, b). Also the leaf areas recorded in plants supplemented with 2–10 μM Se in the selenate form was considerably higher (about 12–27 %) than in the control plants (Fig. 1b).

Effects of increased selenite (a) and selenate (b) concentrations in nutrient solution on the FW and LA of the lettuce plants. Bar SD (n = 24). Mean values marked with the different letters are significantly different (p < 0.05) by Tukey’s multiple comparison test
Under increasing concentrations of selenite the concentration of chlorophyll a tended to decrease, whereas levels of the remaining pigments were little affected (Fig. 2a). The concentrations of photosynthetic pigments have not changed significantly up to selenate concentrations that reached 20 μM (Fig. 1b). When lettuce plants were grown at 20 μM of selenate, the concentrations of all assimilating pigments increased significantly by about 17–19 %, which together with the decline in biomass indicate metabolic perturbations. However, the severe selenate toxicity at a concentration of >30 μM decreased chlorophylls and carotenoids concentration considerably (Fig. 2b).

Effects of increased selenite (a) and selenate (b) concentrations in nutrient solution on the photosynthetic pigments concentrations in the lettuce leaves. Bar SD (n = 6). Mean values for each pigment class marked with the different letters are significantly different (p < 0.05) by Tukey’s multiple comparison test
By comparing values of the toxicity threshold it is evident that selenate is a little less toxic for lettuce cv. Justyna plants than selenite, but this difference is not very great. After passing the toxic threshold concentration, both forms exerted similar adverse effects on plant growth, manifesting in a significant reduction of the roots and shoots biomass, reduction of leaf areas and the concentrations of photosynthetic pigments.
Levels of lipid peroxidation and accumulation of H2O2
Lipid peroxidation has been recognised as a major cause of cellular injury in many biological systems of plant and animal origin. The lipid peroxidation of cell membranes was estimated by measuring the concentration of MDA, which is a by-product of lipid peroxidation in tissue extracts (Largillière and Mélancon 1988). The results indicate that in the leaves of plants biofortified with Se, the concentration of MDA generally increased with respect to control, but a significant increase only at 6 and 15 μM selenite concentrations was noted (Table 1). On the other hand, in the roots a slight reduction of MDA level in the presence of Se was registered at selenite concentrations up to 6 μM and selenate concentrations up to 15 μM. Nonetheless, under highest selenite application (15 μM) there was a significant increase (by about 58 %) in the MDA concentration in the roots.
In addition, Table 1 shows a progressive decrease in the foliar H2O2 concentration as the selenate application rate increases, but a significantly lower (by 27 %) level of H2O2 was recorded only at 15 μM concentration of this Se form. When selenite was applied to the nutrient solution, the foliar H2O2 concentration was slightly reduced with respect to control, but these differences did not reach a statistical significance. In relation to H2O2 concentration in the roots, the results indicate that there were no significant differences in its level according to the Se application rate, except for a significant increase of H2O2 (by about 40 %) in plants supplied with 15 μM of selenite.
Se accumulation and translocation
As expected, the Se in the roots and shoots of lettuce plants increased by increasing the Se concentration in the nutrient solution (Table 2). The obtained results show that the Se concentration in the plant’s organs depended on the rate and form of Se applied to the medium. The Se concentration in roots was much greater after the application of selenite in comparison to the selenate, both forms presenting maximum values after plant exposure to 15 μM Se. In the edible parts of lettuce plants, the Se level was higher in selenate presence compared to the selenite.
In general, Se transport from root to shoot was highly dependent on the chemical form of the Se supplied. The TF value (shoot/root Se concentration ratio) ranged from 0.69 to 2.47 for plants supplied with selenate and was less than 0.15 for those supplied with selenite (Table 2). Maximum translocation’s effectiveness was attained when Se was added as selenate at the concentration of 2 μM, where the TF reached a value of about 2.5. Moreover, if the selenite dose increased in the nutrient solution the TF has tended to increase too.
Total S concentrations
The total S concentration in shoots (Table 3) tended to decrease in response to selenite treatment, but a significantly lower S concentration only in 15 μM selenite treated plants was noted. The opposite pattern was observed on the S level in response to selenate treatment, where S accumulation was over 2-fold higher in plants exposed to 15 μM of selenate compared to control plants. In the roots of plants growing in Se presence, both as selenite and selenate, the S concentration generally was lower than that in the control plants (Table 3). With the exception of plants exposed to 2 μM of selenite, the S concentration in the roots was reduced by 8–22 % in relation to the control value.
Discussion
Although there is increasing evidence that Se enrichment of plants can exert beneficial effects on their growth, antioxidant capacity and stress tolerance (Hartikainen 2005), Se however has not been recognised as an essential micronutrient for higher plants (Seppänen et al. 2010). This paper reports the comparative effects of selenite and selenate on the lettuce plants, with a special focus on the use of this species for biofortification with Se. Lettuce plants were grown in nutrient solutions containing various concentrations of selenite or selenate to determine how these two Se forms affect the growth, photosynthetic pigments levels, oxidant parameters and concentrations of Se and S in plants. The results of the present study demonstrated that an addition of Se to the nutrient solution exerted a dual effect on the growth of lettuce plants depending on Se form and concentration. A marked decrease in FW, leaf area, and photosynthetic pigments concentration was found after passing the toxicity threshold, which has been designated at a level of 15 μM for selenite and 20 μM for selenate. By comparing values of the toxicity threshold it is evident that selenate is a little less toxic for lettuce cv. Justyna than selenite; however, this difference is not very meaningful as was noted in lettuce cv. Philipus by Ríos et al. (2009). The observed toxic effect of Se on plants may result mainly from interferences of Se with S metabolism and from replacing the S-amino acids with corresponding Se-amino acids and their subsequent incorporation into proteins (Hajiboland and Amjad 2007). However, at lower concentrations (<10 μM), Se generally exerted beneficial effects on the plant’s growth. While biofortification of lettuce with Se (2–10 μM) did not affect the accumulation of assimilation pigments, it had a positive effect on the plant’s growth, as the biomass of roots and/or shoots generally increased after Se supplementation, particularly in the presence of selenite. In addition, the leaf areas recorded in plants supplemented with 2–10 μM Se as selenate was considerably higher than in the control plants. The dual effect of Se on plant growth (beneficial or toxic) dependent on Se concentration was also observed in lettuce (Ríos et al. 2009; Ramos et al. 2010), ryegrass (Hartikainen et al. 2000) and other plant species (White et al. 2004; Hajiboland and Amjad 2007).
The results obtained in this study agree with those obtained in other works, which reported that increasing doses of Se in medium culture can cause a significant increase of Se concentration in agricultural crops (Broadley et al. 2006; Broadley et al. 2010; Ramos et al. 2010, Seppänen et al. 2010; Chilimba et al. 2012). Se accumulation in above-ground parts of lettuce was greater when selenate rather than selenite was introduced to the nutrient solution. This finding agrees with studies on lettuce (Ríos et al. 2009; Ramos et al. 2010) and other plant species (Zayed et al. 1998; Longchamp et al. 2012), where Se-enrichment with selenate proved to be more efficient in increasing the Se concentration in shoots than that with selenite. Moreover, the translocation of Se was more efficient when Se was supplied as selenate. When the plants received the lowest dose of Se (2 μM), approximately 70 % of Se taken up when plants were supplied selenate and only 7 % taken up when plants were supplied selenite were found in the shoots. Selenate is more easily transported from the roots and accumulated in the shoots than either selenite or organic Se, which is because much of the selenite is retained in the roots where it is rapidly converted into organic forms, particularly SeMet (Zayed et al. 1998).
Shoot Se concentration in biofortified plants (under non-toxic Se concentrations) varied from 3.7 to 11.3 mg kg−1 DW in selenite treated plants and from 4.7 to 43.3 mg kg−1 DW in selenate treated plants. The recommended consumption of Se for adult men and women is 55–70 μg day−1 (National Academy of Science Dietary Reference 2000). Considering that the average consumption of fresh lettuce by European Union citizens is 23.6 g day−1 (EFSA 2011) and lettuce contains about 95 % of water, the Se concentrations obtained it this study do not cover the daily requirement for Se, but can significantly improve the Se status in humans, especially if the Se concentration in nutrient solution is >2 μM.
One of the causes of lipid peroxidation is H2O2 accumulation which, like other ROS, attacks lipid membranes, thereby increasing the level of MDA (Djanaguiraman et al. 2005). In this study Se application generally did not induce significant changes in lipid peroxidation, with the exception of an increase in MDA level under toxic concentration of selenite (15 μM). Ríos et al. (2009) demonstrated that a concentration of selenite greater than 10 μM provoked a decline in the shoot biomass of lettuce, associated with a decrease in the enzymes that detoxify H2O2, which provoked a rise in lipid peroxidation. However, an application of selenate to a level of 80 μM did not cause an increase in the accumulation of H2O2 in the leaves. In the present study a progressive decrease in foliar H2O2 concentration was noted as the selenate application rate increased from 2 to 15 μM. When selenite was applied to the nutrient solution, the foliar H2O2 concentration was slightly reduced, but not significantly different from controls. In roots there were no significant differences in the level of H2O2 under Se treatment, except for an increase of the H2O2 in 15 μM selenite exposed plants. The increased accumulation of H2O2 and MDA in 15 μM selenite supplied lettuce suggests the occurrence of oxidative stress and could partially explain the toxic effects of Se. Similar results were reported in lettuce cv. Vera by Ramos et al. (2010) and lettuce cv. Philipus by Ríos et al. (2009).
In plants, Se and S metabolism are closely interrelated (Terry et al. 2000); therefore, in the biofortified lettuce the total S accumulation was determined. These results imply that in the above-ground parts of plants there was no significant relationship between these two elements at concentrations of less than 15 μM Se, but at a concentration of 15 μM Se, the S accumulation significantly increased in selenate supplied plants and decreased in selenite supplied plants. In the roots of plants growing in Se presence, both as selenite and selenate, the total S concentration generally was lower than that in the control plants. These results suggest that the presence of Se in the growth medium, particularly at 15 μM concentration, can affect the accumulation and distribution of S between lettuce roots and shoots in different way for selenite and selenate. These data are consistent with previous observations that increasing selenate concentrations in the root area increases S concentrations in shoots of many plant species (Feist and Parker 2001; Suarez et al. 2003; White et al. 2004; Ríos et al. 2008). The interactions between Se and S nutrition were investigated in the model plant Arabidopsis thaliana by White et al. (2004). The results of their study suggest that exogenous selenate can promote sulphate accumulation in the shoots, possibly by preventing a reduction in the abundance and/or activity of sulphate transporters by sulphate and its metabolites. The results of Ríos et al. (2008) indicated that in lettuce plants exposed to 40–120 μM of selenate, the total foliar S concentration increased significantly but the application of selenite did not affect the foliar concentration of S. However, in the present study the toxic concentration of selenite (15 μM) affected S accumulation in lettuce shoots by decreasing the concentration of this macronutrient.
Conclusion
In this study, the effects of two Se inorganic compounds (selenite and selenate) on hydroponic grown lettuce plants were compared, with special attention to biofortification of this species with Se. These results demonstrated that selenite and selenate introduced to the nutrient solution at a concentration below 15 μM can significantly increases the Se concentration in the edible parts of lettuce plants without a negative influence on growth, concentration of photosynthetic pigments, oxidant status as well as S accumulation. Moreover, the growth-promoting effect of Se was observed in Se-biofortified plants. In summary, it is important to emphasise that the biofortification of hydroponic growing lettuce with Se allows an attractive and easy opportunity for increasing the Se concentration in human diets.
Abbreviations
- BHT:
-
Butylated hydroxytoulene
- DW:
-
Dry weight
- FW:
-
Fresh weight
- MDA:
-
Malondialdehyde
- SeCys:
-
Selenocysteine
- SeMet:
-
Selenomethionine
- TBA:
-
Thiobarbituric acid
- TCA:
-
Trichloroacetic acid
- TF:
-
Translocation factor
References
Broadley MR, White PJ, Bryson RJ, Meacham MC, Bowen HC, Johnson SE, Hawkesford MJ, McGrath SP, Zhao FJ, Breward N, Harriman M, Tucker M (2006) Biofortification of UK food crops with selenium. Proc Nutr Soc 65:169–181
Broadley MR, Alcock J, Alford J, Cartwright P, Foot I, Fairweather-Tait SJ, Hart DJ, Hurst R, Knott P, McGrath SP, Meacham MC, Norman K, Mowat H, Scott P, Stroud JL, Tovey M, Tucker M, White PJ, Young SD, Zhao FJ (2010) Selenium biofortification of high-yielding winter wheat (Triticum aestivum L.) by liquid or granular Se fertilization. Plant Soil 332:5–18
Broadley MR, Brown P, Cakmak I, Feng Ma J, Rengel Z, Zhao F (2012) Beneficial elements. In: Marschner P (ed) Mineral nutrition of higher plants, 3rd edn. Academic Press, London, pp 249–269
Bryszewska MA, Ambroziak W, Langford NJ, Baxter MJ, Colyer A, Lewis JD (2007) The effect of consumption of selenium enriched rye/wheat sourdough bread on the body’s selenium status. Plant Foods Hum Nutr 62:121–126
Chilimba ADC, Younga SD, Blacka CR, Meachama M, Lammelc J, Broadley MR (2012) Agronomic biofortification of maize with selenium (Se) in Malawi. Field Crop Res 125:118–128
Dębski B, Zachara B, Wąsowicz W (2001) An attempt to evaluate the level of selenium in Poland and its influence on the healthiness of people and animals. Folia Univ Agric Stetin Zootech 224:31–38 [in Polish]
Djanaguiraman M, Devi DD, Shanker AK, Sheeba JA, Bangarusamy U (2005) Selenium—an antioxidative protectant in soybean during senescence. Plant Soil 272:77–86
European Food Safety Authority (EFSA) (2011) Urgent advice on the public health risk of Shiga-toxin producing Escherichia coli in fresh vegetables. Available at http://www.efsa.europa.eu/en/efsajournal/doc/2274.pdf. Accessed 26 July 2012
Feist LJ, Parker DR (2001) Ecotypic variation in selenium accumulation among populations of Stanleya pinnata. New Phytol 149:61–69
Garcia-Banuelos ML, Hermosillo-Cereceres MA, Sanchez E (2011) The importance of selenium biofortification in food crops. Curr Nutr Food Sci 7:181–190
Hajiboland R, Amjad L (2007) Does antioxidant capacity of leaves play a role in growth response to selenium at different sulfur nutritional status? Plant Soil Environ 53:207–215
Hartikainen H (2005) Biogeochemistry of selenium and its impact on food chain quality and human health. J Trace Elem Med Biol 18:309–318
Hartikainen H, Xue T, Piironen V (2000) Selenium as an anti-oxidant and pro-oxidant in ryegrass. Plant Soil 225:193–200
Haug A, Graham RD, Christophersen OA, Lyons GH (2007) How to use the world’s scarce selenium resources efficiently to increase the selenium concentration in food. Microb Ecol Health Dis 19:209–228
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198
Hoagland DR, Arnon DI (1950) The water-culture method for growing plants without soil. Calif Agric Exp Stn Circ 347:1–32
Jana S, Choudhuri MA (1982) Glycolate metabolism of three submerged aquatic angiosperms during aging. Aquat Bot 12:345–354
Largillière C, Mélancon SB (1988) Free malondialdehyde determination in human plasma by high-performance liquid chromatography. Anal Biochem 170:123–126
Li Z, Zhao X, Sandhu AK, Gu L (2010) Effects of exogenous abscisic acid on yield, antioxidant capacities, and phytochemical contents of greenhouse grown lettuces. J Agric Food Chem 58:6503–6509
Lichtenthaler HK, Wellburn AR (1983) Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem Soc Trans 603:591–592
Longchamp M, Angeli N, Castrec-Rouelle M (2012) Selenium uptake in Zea mays supplied with selenate or selenite under hydroponic conditions. Plant Soil. doi:10.1007/s11104-012-1259-7
Lyons GH, Genc Y, Soole K, Stangoulis JCR, Liu F, Graham RD (2009) Selenium increases seed production in Brassica. Plant Soil 318:73–80
National Academy of Science Dietary Reference (2000) Intakes for vitamin C, vitamin E, selenium and carotenoids. National Academy Press, Washington, DC
Oldfield JE (2002) Selenium word atlas: updated edition. Selenium-Tellurium Development Association, Grimbergen
Ramos SJ, Faquin V, Guilherme LRG, Castro EM, Ávila FW, Carvalho GS, Bastos CEA, Oliveira C (2010) Selenium biofortification and antioxidant activity in lettuce plants fed with selenate and selenite. Plant Soil Environ 56:584–588
Rayman MP (2000) The importance of selenium to human health. Lancet 356:233–241
Ríos JJ, Blasco B, Cervilla LM, Rubio-Wilhelmi MM, Ruiz JM, Romero L (2008) Regulation of sulphur assimilation in lettuce plants in the presence of selenium. Plant Growth Regul 56:43–51
Ríos JJ, Blasco B, Cervilla LM, Rosales MA, Sanchez-Rodriguez E, Romero L, Ruiz JM (2009) Production and detoxification of H2O2 in lettuce plants exposed to selenium. Ann Appl Biol 154:107–116
Seppänen MM, Kontturi J, Heras IL, Madrid Y, Cámara C, Hartikainen H (2010) Agronomic biofortification of Brassica with selenium: enrichment of SeMet and its identification in Brassica seeds and meal. Plant Soil 337:273–283
Suarez DL, Grieve CM, Poss JA (2003) Irrigation method affects selenium accumulation in forage Brassica species. J Plant Nutr 26:191–201
Terry N, Zayed AM, De Souza MP, Tarun AS (2000) Selenium in higher plants. Ann Rev Plant Physiol Plant Mol Biol 51:401–432
White PJ, Broadley MR (2009) Biofortification of crops with seven mineral elements often lacking inhuman diets—iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol 182:49–84
White PJ, Bowen HC, Parmaguru P, Fritz M, Spracklen WP, Spiby RE, Meacham MC, Mead A, Harriman M, Trueman LJ, Smith BM, Thomas B, Broadley MR (2004) Interactions between selenium and sulphur nutrition in Arabidopsis thaliana. J Exp Bot 55:1927–1937
Zachara BA, Pilecki A (2000) Selenium concentration in the milk of breast-feeding mothers and its geographic distribution. Environ Health Persp 108:1043–1046
Zayed A, Lytle CM, Terry N (1998) Accumulation and volatilization of different chemical species of selenium by plants. Planta 206:284–292
Acknowledgments
This research was supported by the Grant No. N N310 430939 of Polish Ministry of Science and Higher Education. Special thanks for technical support and help with the experiments are extended to Monika Kalinowska.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Hawrylak-Nowak, B. Comparative effects of selenite and selenate on growth and selenium accumulation in lettuce plants under hydroponic conditions. Plant Growth Regul 70, 149–157 (2013). https://doi.org/10.1007/s10725-013-9788-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-013-9788-5