
Abstract
This paper is an extension of a previous work which proposes a non-phenomenological model of population growth that is based on the interactions among the individuals of a population. In addition to what had already been studied—that the individuals interact competitively—in the present work it is also considered that the individuals interact cooperatively. As a consequence of this new consideration, a richer dynamics is observed. For instance, besides getting the population models already reached from the original version of the model (as the Malthus, Verhulst, Gompertz, Richards, Bertalanffy and power-law growth models), the new formulation also reaches the von Foerster growth model and also a regime of divergence of the population at a finite time. An agent-based model is also presented in order to give support to the analytical results. Moreover, this new approach of the model explains the Allee effect as an emergent behavior of the cooperative and competitive interactions among the individuals. The Allee effect is the characteristic of some populations of increasing the population growth rate in a small-sized population. Whereas the models presented in the literature explain the Allee effect with phenomenological ideas, the model presented here explains this effect by the interactions between the individuals. The model is tested with empirical data to justify its formulation. Another interesting macroscopic emergent behavior from the model proposed is the observation of a regime of population divergence at a finite time. It is interesting that this characteristic is observed in humanity’s global population growth. It is shown that in a regime of cooperation, the model fits very well to the human population growth data from 1000 AD to nowadays.










Similar content being viewed by others

Notes
This consideration of the interaction field when \(r<r_0\) differs from the MLBI model, but do not change its qualitative aspect. Moreover, it brings more generality to the model.
References
Allee WC et al (1949) Principles of animal ecology. Saunders, London
Arruda T-J, González R-S, Terçariol C-A-S, Martinez A-S (2008) Arithmetical and geometrical means of generalized logarithmic and exponential functions: generalized sum and product operators. Phys Lett A 372:2578
Ausloos Marcel (2012) Another analytic view about quantifying social forces. arXiv:1208.6179, Submitted on 30 (Aug 2012)
Barberis L, Condat CA, Romãn P (2011) Vector growth universalities. Chaos, Solitons & Fractals 44:1100–1105
Bettencourt LMA et al (2007) Growth, innovation, scaling, and the pace of life in cities. PNAS, 104(17):7301–7306
Boccara N (2003) Modeling complex systems (graduate texts in contemporary physics). Springer, Berlin
Cabella BCT, Martinez AS, Ribeiro F (2011) Data collapse, scaling functions, and analytical solutions of generalized growth models. Phys Rev E 83:061902
Cabella BCT, Ribeiro F, Martinez AS (2012) Effective carrying capacity and analytical solution of a particular case of the Richards-like two-species population dynamics model. Phys A 391:1281–1286
Chester M (2011) A law of nature? Open J Ecol 1(3):77–84
Courchamp F, Clutton-Brock T and Grenfell B (1999) Inverse density dependence and the Allee effect. Tree 14(10)
d’Onofrio A. Fractal growth of tumors and other cellular populations: linking the mechanistic to the phenomenological modeling and vice versa. Chaos, Solitons & Fractals (2009). doi:10.1016/j.chaos.2008.04.014
dos Santos LS, Cabella BCT, Martinez AS (2014) Generalized Allee effect model. Theory Biosci 133:117–124
Drasdo D, Hohme S (2003) Individual-based approaches to birth and death in avascular tumors. Math Comput Model 37:1163–1175
Edelstein-Keshet L (2005) Mathematical models in biology (classics in applied mathematics), 1st edn. SIAM: Society for Industrial and Applied Mathematics, Philadelphia
Falconer KJ (1990) Fractal geometry: mathematical foundations and applications. Wiley, New York
Freyer JP, Sutherland RM (1985) A reduction in the in situ rates of oxygen and glucose consumption of cells in EMTG/Ro spheroids during growth. J Cell Physiol 124:516–524
Ghazoul J, Liston KA, Boyle TJB (1998) Disturbance-induced density dependent seed set in Shorea siamensis (Dipterocarpaceae), a tropical forest tree. J Ecol 86:462–473
Ginzburg LR (1972) The analogies of the “free motion” and “force” concepts in population theory. In: Ratner VA (ed) Studies on theoretical genetics. Academy of Sciences of the USSR, Novosibirsk, pp 65–85
Gompertz R (1825) On the nature of the function expressive of the law of human mortality, and on a new mode of determining the value of life contingencies. Philos Trans R Soc Lond 115:513–585
Gregorczyk A (1998) Richards plant growth model. J Agron Crop Sci 181(4):243–247
Gregory SD, Bradshaw CJA, Brook BW, Courchamp FC (2009) Limited evidence for the demographic Allee effect from numerous species across taxa. Ecology 91:2151–2161
Groom MJ (1998) Allee effects limit population viability of an annual plant. Am Nat 151:487–496
Guiot C, Degiorgis PG, Delsanto PP, Gabriele P, Deisboeck TS (2003) Does tumor growth follow a “universal law”? J Theor Biol 225(2):147–151
Haybittle JL (1998) The use of the Gompertz function to relate changes in life expectancy to the standardized mortality ratio. Int J Epidemiol 27(5):885–889
Kadanoff LP (2000) Statistical physics: statics,dynamics and remormalization. World Scientific, Singapore
Kohler TA, Gumerman GG (2000) Dynamics in human and primate societies. Oxford University Press, Oxford
Kuehn C, Siegmund S, Gross T (2011) On the dynamical analysis of evolution equations via generalized models. arXiv:1012.4340
Malthus TR (1798) An essay on the principle of population as it affects the future improvement of society. J. Johnson, London
Martinez A-S, González R-S, Terçariol C-A-S (2008) Continuous growth models in terms of generalized logarithm and exponential functions. Phys A 387:5679
Martinez A-S, González R-S, Espíndola A-L (2009) Generalized exponential function and discrete growth models. Phys A 388:2922
Mitchell M (2011) Complexity: a guided tour. Oxford University Press, Oxford
Mombach JCM, Lemke N, Bodmann BEJ, Idiart MAP (2002) A mean-field theory of cellular growth. Europhys Lett 59(6):923–928
Murray JD (2002) Mathematical biology I: an introduction. Springer, New York
Ribeiro F, Cabella BCT, Martinez AS (2014) Richards-like two species population dynamics model. Theory Biosci 133:135–143. doi:10.1007/s12064-014-0205-z
Richards FJ (1959) A flexible growth function for empirical use. J Exp Bot 10(2):290–301. doi:10.1093/jxb/10.2.290
Roll J et al (1997) Reproductive success increases with local density of conspecifics in a desert mustard (Lesquerella fendleri). Conserv Biol 11:738–746
Savageau MA (1980) Growth equations: a general equation and a survey of special cases. Math Biosci 48:267–278
Sibly RM, Barker D, Denham MC, Hone J, Pagel M (2005) On the Regulation of populations of mammals, birds, fish, and insects. Science 309
Solomon S (1999) Generalized Lotka-Volterra (GLV) models and generic emergence of scaling laws in stock markets. arXiv:cond-mat/9901250
Strzalka D (2009) Connections between von Foerster coalition growth model and Tsallis q-exponential. Acta Physica Polonica B, 40(1)
Strzalka D, Grabowski F (2008) Towards possible q-generalizations of the Malthus and Verhulst growth models. Physica A 387(11):2511–2518
Tsallis C (1988) Possible generalization of Boltzmann-Gibbs statistics. J Stat Phys 52:479
Tsallis C (1994) What are the numbers that experiments provide? J Química Nova 17:468
Verhulst PF (1845) Recherches mathematiques sur la loi d’accroissement de la population. Nouveaux memoires de l’Academie Royale des Sciences et Belles Lettres de Bruxelles 18:1–38
Verhulst PF (1847) Deuxieme memoire sur la loi d’accroissement de la pop- ulation. Nouveaux memoires de l’Academie Royale des Sciences et Belles Lettres de Bruxelles 20:1–32
von Bertalanffy L (1957) Quantitative laws in metabolism and growth. Q Rev Biol 32(3):217–231
von Bertalanffy L (1960) Principles and theory of growth. In: Nowinski WW (ed) Fundamental aspects of normal and malignant growth. Elsevier, New York, pp 137–259
Von Foerster H, Mora PM, Amiot WL (1960) Doomsday: Friday, 13 november, AD 2026 at this date human population will approach infinite if it grows as it has grown in the last two millenia. Science 132
West GB, Brown JH, Enquist BJ (2001) A general model for ontogenetic growth. Nature, 413:628–631
West GB et al (1997) A general model for the origin of allometric scaling laws in biology. Science 276:122. doi:10.1126/science.276.5309.122
Yeomans JM (1992) Statistical mechanics of phase transitions. Oxford University Press, Oxford
Acknowledgments
I would like to acknowledge the useful and stimulating discussions with Alexandre Souto Martinez and Brenno Troca Cabella. I would like to acknowledge also the support from CNPq (151057/2009-5).
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix 1: The Generalized Logarithm and Exponential Function
In this appendix, one presents the generalizations of the logarithmic and exponential functions and some of their properties. The introduction of the functions is shown to be very useful for dealing with the mathematical representation of the population growth model that is presented in this work.
The \(\tilde{q}\)-logarithm function is defined as
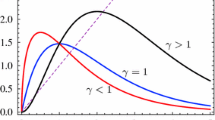
which is the area of the crooked hyperbole and is controlled by \(\tilde{q}\). This equation is a generalization of the natural logarithm function, which is reproduced when \(\tilde{q} = 0\). This function was introduced in the context of nonextensive statistical mechanics Tsallis (1988; 1994) and was studied recently by Arruda et al. (2008), Martinez et al. (2008) and Martinez et al. (2009). Some of the properties of this function are as follows: for \(\tilde{q} < 0, \ln _{\tilde{q}}(\infty )=-1/\tilde{q}\); for \(\tilde{q} > 0, \ln _{\tilde{q}}(0)=-1/\tilde{q}\); for all \(\tilde{q}, \ln _{\tilde{q}}(1)=0\); \(\ln _{\tilde{q}}(x^{-1}) = - \ln _{-\tilde{q}}(x)\); \(\hbox {d} \ln _{\tilde{q}}(x)/\hbox {d}x = x^{\tilde{q}-1}\). Moreover, the \(\tilde{q}\)-logarithm is a function: convex for \(\tilde{q}>1\), linear for \(\tilde{q}=1\), and concave for \(\tilde{q}<1\).
The inverse of the \(\tilde{q}\)-logarithm function is the \(\tilde{q}\)-exponential function, which is by
Some properties of this function are as follows: \(\hbox {e}_{\tilde{q}}(0)=1\), for all \(\tilde{q}\); \( \left[ \hbox {e}_{\tilde{q}}(x) \right] ^a = \hbox {e}_{\tilde{q}/a}(ax)\), where \(a\) is a constant; for \(a=-1\), one has \(1/\hbox {e}_{\tilde{q}}(x) = \hbox {e}_{-\tilde{q}}(-x)\). Moreover, the \(\tilde{q}\)-exponential is a function: convex for \(\tilde{q}<1\); linear for \(\tilde{q}=1\); concave for \(\tilde{q}>1\).
Appendix 2: A Detailed Calculus of \(I_i^{(l)}\)
In this appendix, one presents a detailed calculus for the intensity of the interaction felt by a single individual \(i\) from the other individuals of the population, which is represented by \(I_i^{(l)}\) (see Sect. 2). One follows Mombach et al. (2002) to show that this intensity is independent of the individual; that is, it is the same for all individuals of the population and depends only on the size of the population. More specifically, one shows that \(I_i^{(l)} = I^{(l)}(N)\) regardless of \(i\).
First, from the Sect. 2 one has
where \(\delta _{ij}\), which is the Kronecker’s delta, was introduced to avoid the restriction in the sum. Moreover, \(\mathbf {r}_i\) and \(\mathbf {r}_j\) represent the position vectors of the individuals \(i\) and \(j\), respectively, and consequently \(r_{ij} = |\mathbf {r}_i - \mathbf {r}_j |\) is the distance between them. Introducing the property
where \(\delta (\cdots )\) is the Dirac’s delta, the expression (35) becomes
In the last two expressions, was introduced: \(D (=1,2, 3)\), which is the Euclidean dimension in which the population is embedded, and \(V_{D}\), which is the total (hyper) volume (in \(D\) dimensions) that contains the population. The form represented in (37) was obtained by the variable substitution \(\mathbf {r}_j - \mathbf {r}_i\) by \(\mathbf {r}\), using Dirac’s delta.
Some algebraic manipulation and the introduction of \(r\equiv |\mathbf {r}|\) allows to write
Note that \(dN(\mathbf {r})\equiv \hbox {d}^D \mathbf {r} \sum _{j \ne i} \delta \Big ( \mathbf {r} - (\mathbf {r}_j - \mathbf {r}_i)\Big )\) is the number of individuals which is at the element of (hipper)volume \(d^D\mathbf {r}\) at the distance \(\mathbf {r}\) from the individual \(i\), localized at \(\mathbf {r}_i\). In this way, the density of individuals at \(\mathbf {r}_i + \mathbf {r}\) (neighbors of \(i\)), that is \(\rho (\mathbf {r}_i~+~\mathbf {r}) =\hbox {d}N(\mathbf {r})/\hbox {d}^D \mathbf {r}\), can be written as
The density of individuals can also be thought of in terms of the scale of the system (in conformity with Falconer 1990). The volume of the system grows in the form \(V_{D} \sim L^D\), where \(L\) is the typical size of the system. However, the number of individuals grows as the form \(N \sim L^{D_{f}}\), where \(D_{f}\) is the fractal dimension formed by the spatial structure of the population. By considering \(r\), which is the absolute distance from \(i\), as a typical distance of the system, one can say that the density of individuals (\(V_{D}/N\)) has the form
where \(\rho _0\) is a constant which is related to the density of individuals. In fact, if \(D=D_{f}\) and the population is homogeneously distributed, then \(\rho _0\) is the usual density of individuals.
Using results (40) and (39) in (38), one obtains
Note that the integration argument does not depend on the angular coordinates. Thus, one can write \(\hbox {d}^D \mathbf {r}=~r^{D-1}\hbox {d}r \hbox {d}\Omega _{D}\), where \(\hbox {d}\Omega _{D}\) is the solid angle, which implies
where \(\Omega _{D} = \int \hbox {d}\Omega _{D}\). Note that the only term that depends on the Euclidean dimension is the solid angle, and \(\Omega _{D}\) assumes the following values according to these tree possibilities: \(\Omega _1 = 2\); \(\Omega _2 = 2\pi \); \(\Omega _3 = 4\pi \). By introducing the constant \(\omega _{D} = \rho _0 \Omega _{D}\), which depends only on \(D\), one obtains
Thus, \(I_i^{(l)}\) does not depend on the label \(i\) anymore. As a result, one can say that \(I_i^{(l)} = I^{(l)}\) regardless of \(i\).
Furthermore, one can introduce the total number of individuals in the relation above by the following thinking. The total number of individuals in the population can be determined by the integral
Using Eq. (40) and integrating the solid angle, one obtains
Note that the first term on the right in (46) and (47) can be zero (indicating the absence of individuals) or 1 (indicating the presence of a single individual). These values are possible because the ratio of the individual is \(r_0\), and hence, there can be at most one individual inside the region that consists of the length between \(0\) and \(r_0\). Thus, for \(r_0=1, \omega _d/D_{f} \sim 1\). \(R_{\hbox {max}}\) can be obtained from (47), which is a function of \(N\) according to
Returning to relation (43), one finds
By introducing \(\tilde{q}_l=1- \gamma _l/D_{f}\) and the properties of the generalized logarithm (Appendix 1), one obtains
Rights and permissions
About this article

Cite this article
Ribeiro, F.L. A Non-phenomenological Model of Competition and Cooperation to Explain Population Growth Behaviors. Bull Math Biol 77, 409–433 (2015). https://doi.org/10.1007/s11538-014-0059-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11538-014-0059-z