
Abstract
Cooperation is ubiquitous ranging from multicellular organisms to human societies. Population structures indicating individuals' limited interaction ranges are crucial to understand this issue. But it remains unknown to what extend multiple interactions involving nonlinearity in payoff influence the cooperation in structured populations. Here we show a rule, which determines the emergence and stabilization of cooperation, under multiple discounted, linear and synergistic interactions. The rule is validated by simulations in homogenous and heterogenous structured populations. We find that the more neighbours there are the harder for cooperation to evolve for multiple interactions with linearity and discounting. For synergistic scenario, however, distinct from its pairwise counterpart, moderate number of neighbours can be the worst, indicating that synergistic interactions work with strangers but not with neighbours. Our results suggest that the combination of different factors which promotes cooperation alone can be worse than that with every single factor.
Similar content being viewed by others

Introduction
The particulars of why and how cooperation evolves have perplexed evolutionary biologists and sociologists enduringly1,2,3,4. A cooperator takes an altruistic action which supplies a benefit, b, for another individual at a cost, c, while a defector does nothing. One of the main tasks of evolutionary theory is to explain why and how cooperation is present. Evolutionary game theory provides a powerful platform to understand the evolution of cooperation in unstructured populations, with the replicator equation in infinite populations3 and stochastic dynamics in finite populations5,6,7. Recently the assumption of a well mixed population is removed and the population allows individuals to interact locally8,9. Typically networks are adopted to depict such population structure, since it is simple in definition, while complex in property8,10. The nodes of the network represent individuals, while the edges denote connections in between11,12. In this way, a network paves the way to capture the intrinsic idea of local interaction13,14: Individuals interact with their neighbours only. In particular, the degree of a node represents the number of neighbours of the focal individual, which indicates the interaction range. These network structures are widespread in human organizations15,16, scientific collaboration among researchers17 and even somatic evolution within multicellular organisms18. However, for structured populations, in contrast with that in the well mixed case5,6, it becomes challenging to analyze the evolutionary dynamics theoretically. This is because enormous possible topological configurations arise during the process of evolution19,20,21. In spite of being challenging, there are advances in the analytical methods12,22,23,24,25,26. The main result is that local interactions can pave the way for the emergence of cooperation.
The conflict between cooperation and defection is captured by the prisoner's dilemma in the beginning2,27, a pairwise game. Though the “Tragedy of Commons”, a multi-player game, was introduced to depict this dilemma long before28, it has not been popular until recently owing to its complexity29,30,31,32,33,34,35,36,37,38. This is also true in structured populations: For pairwise interactions, conditions for cooperators to be selected over defectors have been theoretically investigated in general structured populations12,22,24. For multiple interactions, however, only two extreme types of network structure with cycle39 and well mixed populations32,34,35,40 have been addressed. For the network degree in between the minimum (the cycle) and the maximum (well-mixed population), it is unclear under what conditions cooperation outperforms defection. Besides, we introduce nonlinearity in the public goods, which is intrinsic to the fitness of multi-player games. For simplicity, the synergistic and discountable effects of the public goods are adopted: These effects are wide spread in microbes41,42,43. As a cluster of microbes secretes enzymes to digest the extracellular resource, the benefit of the secreted enzyme (public goods) provided by the first cooperative cell may play a vital role for survival, while the enzymes will eventually be saturated for the resource with the increase of cooperators, thus the cooperator cells joining the group later only contribute diminishing small benefits to the group44. This is the discounting effect of the public goods. While for synergy, enzyme-mediated reactions will be launched by enzyme-producing cooperators. With the concentration of enzyme production, this may exhibit a faster efficiency than linear increase45. In addition, the framework of synergy and discounting effects provides a unifying framework reconciling different social dilemmas41, thus it does not lose generality in spite of its simplicity. The main result in the well mixed population is that synergy is beneficial for the emergence of cooperation while the discounting effect is detrimental.
Considering the importance of both the population structure and the multiple interactions on the evolution of cooperation, we theoretically explore how the combination of these two effects affects the emergence and stabilization of cooperation. To this end, we are addressing the stochastic dynamics of the public goods game with synergy and discounting in a generally random regular graph with arbitrary degree. We find a rule theoretically elucidating the critical value of benefit-to-cost ratio b/c up to which cooperation emerges and is stabilized. In addition numerical simulations verify the validity of the rule as well as its feasibility for random graph. For linear public goods game on any regular graph and any public goods game with synergy and discounting on a cycle, we find an equivalent proposition that the rule determines not only the emergence, but also the stabilization of cooperation. What's more, in public goods with synergy, we present that it can be the worst for the emergence of cooperation, as the number of neighbours is moderate. We find that synergistic interactions work with strangers but not with neighbours and cooperation with both synergistic and local interactions can be worse than that with each alone. Our work suggests that there can be a big shadow in the effects of combinational mechanisms on the evolution of cooperation.
Results
Description of the model
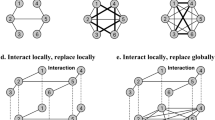
We consider a finite population located on a graph of size N. Individuals are assigned to the nodes of the graph, whereas social ties between them are represented by the edges11,12. Every individual has k neighbours. As illustrated in Fig. 1, players participate in the public goods game organized by themselves and their neighbours46, that is to say, each player participates in k + 1 public goods games of size n = k + 1.

Illustration of updating on a network.
We show a network with size N = 22 and every player has k = 3 neighbours here. Cooperator C11 and defector D12 (D13) are neighbours of the selected cooperator C1 with updating. Both of C11 and D12 (D13) have neighbours with strategy of cooperation or defection except C1, which are called C111 and D112, C121 (C131) and D122 (D132), respectively. C1a1 and D1b2 also have neighbours C1a11, C1b21 adopting strategy of cooperation and D1a12, D1b22 with defection, where both of a and b mean 1, 2, or 3. Each player organizes a public goods game with all of its k neighbours. Thus each individual participates in k + 1 public goods games of size k + 146. As an example, for the payoff of C11, all players marked within the dashed curve are relevant. The payoff comes from all games C11 participates in, where one game (shaded in blue) held by C11, that is, part (a) and the other three (shaded in red) held by C1, C111 and D112, that is, part (b), (c) and (d).
For the public goods game, the first cooperator contributes a benefit b while the jth (1 ≤ j ≤ n) cooperator contributes bδj−1 to the common pool41. Every cooperator pays the same cost c. Defectors exploit the group by reaping benefits without paying anything. The accumulated benefits are distributed equally to all the n players in the group irrespective of their behaviors. Thus, defectors and cooperators receive the following payoffs41


where i is the number of cooperators within the group. Here δ > 0 is regarded as the discounting (0 < δ < 1) or synergy (δ > 1) factor. As δ = 1, it degenerates to the linear public goods game with PD(i) = rci/n, where r = b/c is the multiplication factor.
After playing the public goods game, the payoff P of every player is transformed into fitness f by fitness mapping5,47. Here we adopt the linear fitness which consists of baseline fitness and the payoffs arising from games5,12, i.e., f = 1 − w + wP where w varying from 0 to 1 is the intensity of selection. For w → 0, the selection is weak. It means that the game is merely one of many factors which contribute to the entire fitness of an individual5,12.
As to the updating rule, the “death-birth” (DB) process4 is employed. Within the process, a player in a population is randomly selected to die at each time step and then all neighbours of the focused player, with probability proportional to their individual fitness, compete for the vacant site.
A rule for the evolution of cooperation
We study the emergence of cooperation by comparing the fixation probability5,12,37,48 of a single cooperator (ρC) invading a wild population of defective type under weak selection with that under neutrality 1/N4. If ρC > 1/N then natural selection favors cooperator replacing defector4, so we see that natural selection favors the emergence of cooperation. We see that natural selection favors the stabilization of cooperation if ρD < 1/N, that is, natural selection opposes the fixation of defectors. And if ρC > ρD, we see that natural selection favors cooperator over defector4.
We obtain the fixation probability of both cooperation and defection by the pair approximation (Supplementary Notes 1 and 2). For large population size and weak selection, we have a rule:  if and only if
if and only if

and  if and only if
if and only if

where the notations Q1, f(δ) and g(δ) can be found in Methods.
For the linear public goods game (δ = 1), natural selection favors not only the emergence of cooperation but also its stabilization if and only if

On this occasion, the rule is equivalent to r > n2/(n + 2) where r is the multiplication factor of the common pool. Since n2/(n + 2) < n, it implies theoretically that cooperative dilemma can be relaxed in structured populations compared with that in the well mixed case, without invoking any other additional mechanisms. For the linear public goods game on the cycle (δ = 1 and k = 2), ρC > ρD is equivalent to b/c > 9/5, which coincides with39 (Supplementary Note 2.4).
What's more is that the general rule to determine the emergence of cooperation is found to be in good agreement with computer numerical simulations (the first row of Supplementary Fig. S1). And it also approximately applies to heterogeneous structured populations (the second row of Supplementary Fig. S1).
Based on equation (5), we find that ρC > ρD is also equivalent with the emergence ρC > 1/N and stabilization ρD < 1/N of cooperation. Furthermore, the following equivalence holds

for the public goods game which is either linear δ = 1 or on a cycle k = 2. That is to say, for large structured population under linear public goods game or public goods game with nonlinearity in individual payoff on cycle, we have that natural selection favors emergence of cooperation if and only if it favors stabilization of cooperation. Further, we show that the critical value, both for the emergence and the stabilization of cooperation, is continuous with the discounting or synergy factor δ (Supplementary Note 3.1). Hence the equivalent proposition (6) applies for infinitesimal nonlinearity (Supplementary Note 3.2).
Nonlinearity on the evolution of cooperation
From the rule (inequalities (3) and (4) in Methods), we theoretically get the critical benefit-to-cost ratio b/c for the emergence and stabilization of cooperation (defection) with the two factors combined, saying the spatial reciprocity and nonlinearity in payoff induced by multiple interactions. Fig. 2 shows that weak discounting significantly inhibits both the emergence and stabilization of cooperation, whereas the weak synergy favors them greatly. In contrast with the linear public goods game, the critical ratios b/c for ρC = 1/N and ρD = 1/N are no longer overlapping for nonlinear payoff effects (Fig. 2). In other words, taking into account either discounting or synergy, a particular form of nonlinearity in payoff, the emergence and the stabilization of cooperation are no longer equivalent as in the linear public goods case (equivalent proposition (6)).

The critical value of the benefit-to-cost ratio for natural selection favoring emergence and stabilization of cooperation (defection).
We set δ = 0.95 (a) and 1.05 (b) to represent respectively the weak discounting and synergy in structured populations with N = 5 × 103 numerically. For discounting effect δ < 1, both the critical values for ρC = 1/N and ρD = 1/N increase rapidly with average degree k. Yet the critical value for ρD = 1/N is greater than that of ρC = 1/N and it also increases much faster. This shows that in the discounting public goods game, with the increase in the number of neighbours, it is easier for a cooperator to be invasive (ρC > 1/N) than to be stabilized (ρD < 1/N). For synergy effect δ > 1, the critical value for ρC = 1/N is greater than that of ρD = 1/N for any neighbour size. This shows that it is easier for cooperation to be stabilized (ρD < 1/N) than to be invasive (ρC > 1/N). Interestingly, both the critical values for ρC = 1/N and ρD = 1/N are non-monotonic and is a one-hump function of k for weak synergy. This shows for both the emergence and the stabilization of cooperation in the public goods game with weak synergy, a moderate number of neighbours is the worst. Further, for the strong synergy case, we will see that the one-hump degenerates to a decreasing function of k.
For multiple interactions such as linear public goods, similar to pairwise interactions12, cooperation will also be impeded with an increase of the number of neighbours (light gray up triangle in Fig. 2). Discounting in payoff significantly inhibits cooperation (Fig. 2a). In particular, in this case, with the increase in the size of neighbourhood, it will become even harder for the emergence and stabilization of cooperation. The critical benefit-to-cost ratio is increasing much rapidly than its linear public goods game counterpart.
For weak synergy, the critical benefit-to-cost ratio b/c for emergence or stabilization of cooperation still first increases as the growth of every player's number of neighbours (Fig. 2b). But it decreases as neighbour size is big enough and tends towards zero. This illustrates that for small size of neighbour, increasing interaction range, i.e. k, is detrimental for cooperation, which is consistent with linear public goods game; yet the interesting story comes along when the interaction range is relatively large, in this case, increasing the interaction range is beneficial for cooperation, which is seldom observed in cooperative dilemmas. An intuitive understanding can be: for small neighbour size, the local competition plays a key role. Even though the public goods are exponentially increasing with additional cooperator in a group, the group is small in size generally, thus the defector would reproduce more efficiently with the increase of size. This leads to the increase of the critical benefit-to-cost ratio. For large size of neighbours, however, the exponential increase in the accumulation of public goods with an additional cooperator outperforms the reproduction of defectors. For example, with the jth (0 ≤ j ≤ k + 1) cooperator's benefit bδj−1 to common pool, big j induces large payoff bδj−1/(k + 1) to every player in the same group with weak synergistically enhanced effect δ as well as the baseline benefit b. This rather large payoff paves the way for emergence and stabilization of cooperation for large neighbour size. Therefore, it is the worst for emergence of cooperation when the number of neighbours is moderate. Another intuitive explanation is that for weak synergy effect, the replicator dynamics allows the coexistence of cooperators and defectors41, which is quite similar to the snowdrift game. For the snowdrift game, however, local interaction can inhibit cooperation49 in contrast with the prisoner's dilemma50. Here, we explicitly show up to how many numbers would be the worst for cooperation in such scenario. It shows that synergistic interactions work with strangers in well mixed populations41. Furthermore, we also find that the synergistic interactions do not work with neighbours in structured populations.
For stronger synergy, the critical benefit-to-cost ratio always decreases with the neighbour size (Fig. 3). Thus in this case cooperation could be promoted significantly with the increase of every player's number of neighbours under strong synergy, which intrinsically differs from effect of linearity, discounting, or weak synergy. This is because the synergy effect is so great that it effectively is a coexistence game where best relies should be of the population minority. In this case, enlarging the interaction range paves the way for cooperator mutants to be more like an minority yielding an enhancement of cooperation level.

Critical value of benefit-to-cost ratio degenerates to a monotonic function of average degree k from one-hump function with the enhancement of synergy.
We set different synergy factors to investigate its effects on the critical value of b/c with population size N = 5 × 103. Specific values of δ are marked on each panel. Orange star and cyan square are used to indicate ρC = 1/N and ρD = 1/N respectively. The critical value decreases with the increase of δ. And it is a parabolic function of k when synergy is weak ((a), (b), (c), (d) and (e)), whereas monotonic when synergy is strong (f). Under the synergistic effect, big k or δ can drive the critical value to approach 0. The intuitive understanding behind this is the competition between the two factors. For large k, the population is approximately well mixed, thus the local interaction diminishes by the replicator equation41 and cooperation with synergy thrives, leading to a decrease of the critical value with increasing k. For small k, the local interaction plays an important role if the synergy effect is not strong. That is to say for δ slightly more than the unit, increasing k does inhibit the fixation for both strategies as in the pairwise prisoner's dilemma12. To sum up, for small δ > 1, there is a hump for the critical value with the increase of neighbours. For large δ, however, even for small k, the synergy effect is strong enough to outperform the locality of the population structure. By the same argument, the critical value is monotonically decreasing with the neighbours.
Our results show that the intrinsic multiple interactions, where payoffs are nonlinear in general, can lead to a one-hump function of the critical benefit-to-cost ratio to facilitate cooperation, which never happens in pairwise interactions, or even linear multiple interactions. This echoes partly Hamilton's metaphor of novelty of multiple interactions, though he also pointed out that the investigation will be challenging, which is mirrored as sea-sickness51.
Discussion
Over the past two decades, structured populations depicted by networks have been taken into consideration to study the evolution of cooperation by virtue of evolutionary graph theory11,20,24,52. It has been shown that cooperation can flourish in both static network and dynamic network8,12,20,25,52,53 (for an exception, please see49). The main reason is that these population structures can lead to a clustering of cooperative individuals8,12,21,24, within which cooperators can survive by enjoying the benefits from mutual cooperation even though some cooperators are exploited by defectors along cluster boundaries. We indicate that in a non-additive public goods game, where nonlinearity in payoff arises, this clustering (equations (S20) and (S21) in Supplementary Note 1.2) is not always beneficial when the neighbours are few in number: in a synergy public goods scenario, where the latter cooperators in the group contribute significantly much more than the previous cooperators, the worst case for cooperation emerges when the number of neighbours is moderate, not too big nor too small. This means that synergistic interactions work with strangers but not with neighbours. The technical part is challenging for a general multi-player game54. Here we adopt synergistic and discounted effects as an example to indicate the nonlinearity in public goods for mathematically convenience. Synergy and discounting can be used to unify typical social dilemmas such as the prisoner's dilemma and snowdrift game41, thus it also does not lose much of the generality. Our results show that the interaction between different mechanisms55 might trigger novel unexpected results. The combination of different factors with each promoting cooperation alone can be worse than every factor alone in promoting cooperation. Thus, it may be promising to investigate the combination of previous mechanisms promoting cooperation.
We find that the rules governing the emergence and stabilization of cooperation are equivalent for linear public goods games, which is validated by numerical simulations on homogenous as well as heterogenous structured populations. The rule simply asks the benefit to cost ratio b/c to exceed a critical value (k + 1)2/(k + 3), where k is the average number of neighbours in the population.
In fact, for any number of neighbours k, the numerator of the critical value is (k + 1)2, which suggests the number of individuals relevant to the payoff of the focal individual with recounting (Fig. 1), i.e., the product of the group size k + 1 and the average number of the public goods game every player involved in, k + 1. Therefore, in this case, as in a well mixed population, multiple interactions significantly inhibit the cooperation than its pairwise counterpart. For public goods games on a cycle, with either linearity, discounting, or synergy, the equivalence still holds between the rules facilitating the emergence of cooperation and that governing the stabilization. Therefore the same criterion applies to determine under what condition the average abundance of cooperation exceeds that of defection in the mutation-selection equilibrium under small mutation56,57.
The equivalence falls down for general population structures and nonlinear public goods game. For the synergy effect, the emergence and the stabilization of cooperation are facilitated significantly for any number of neighbours compared with the linear public goods game. Being stabilized, in this case, is much easier than the emergence. For the discounting effect, both the emergence and the stabilization of cooperation are inhibited significantly for any number of neighbours compared with the linear public goods game. Being stabilized, in this case, is much harder than the emergence. Therefore, both synergy and discounting has a more significant role in the stabilization compared with the emergence.
We find that our theoretical results can be approximated well by simulations as the synergy or discounting effect is weak, i.e., when δ is close 1 (Supplementary Fig. S1). However, for δ deviating from 1, the pair approximation leads to a large deviation from simulations (Supplementary Fig. S1). The reason for such deviation is mainly in two folds52,58: The first is that pair approximation is formulated for the regular graph without any loops. For our graphs, however, they are not always in this case (Fig. 1); The second is that the approximation is precise for large population size and weak selection, whereas the simulations bear finite population size, which generates the prediction error. In fact, triplet correlations are a more natural and more precise choice for calculating payoffs from group interactions, since one's payoff is not merely related with his or her neighbors but also with the neighbors' neighbors in our model. But the triplet approximation introduces more independent variables, leading to a tedious analysis. Another limitation of our work is the weak selection assumption. In general, results obtained under the weak selection can not be extrapolated to the strong selection directly for multi-player games even in well mixed populations59. Thus we do not expect that our theoretical results stay robust for the strong selection.
As applications, for microbes with either synergy and discounting public goods, in particular, if the public goods are diffusive60, the average degree of the network suggests the diffusion rate of the public goods. Our result suggests that for the discounting public goods game, only a low diffusion rate of public goods can make the cooperator cells thrive; For the synergy public goods game, however, it is always better than the discounting case. The interesting result lies in the fact that cooperation is better off for both very low and very high diffusion, whereas is worst off for moderate diffusion rate. Experimental validation along this line might be interesting.
Methods
Notations in inequalities (3) and (4) are


and



The precise process of the calculation can be found in Supplementary Notes 1 and 2.
Expanding f(δ) and g(δ) in Taylor series around δ = 1 leads to


where o((δ − 1)4) shows the error is the high order infinitesimal of (δ − 1)4. Further analysis can be found in Supplementary Notes 2.4 and 3.
References
Hamilton, W. D. The evolution of altruistic behavior. Am. Nat. 97, 354–356 (1963).
Trivers, R. L. The evolution of reciprocal altruism. Q. Rev. Biol. 46, 35–57 (1971).
Hofbauer, J. & Sigmund, K. Evolutionary Games and Population Dynamics (Cambridge University Press, Cambridge, 1998).
Nowak, M. A. Evolutionary Dynamics: Exploring the Equations of Life (Harvard University Press, Cambridge, MA, 2006).
Nowak, M. A., Sasaki, A., Taylor, C. & Fudenberg, D. Emergence of cooperation and evolutionary stability in finite populations. Nature 428, 646–650 (2004).
Traulsen, A., Claussen, J. C. & Hauert, C. Coevolutionary dynamics: From finite to infinite populations. Phys. Rev. Lett. 95, 238701 (2005).
Traulsen, A., Nowak, M. A. & Pacheco, J. M. Stochastic dynamics of invasion and fixation. Phys. Rev. E 74, 011909 (2006).
Nowak, M. A. & May, R. Evolutionary games and spatial chaos. Nature 359, 826–829 (1992).
Durrett, R. & Levin, S. A. The importance of being discrete (and spatial). Theor. Popul. Biol. 46, 363–394 (1994).
Albert, R. & Barabási, A.-L. Statistical mechanics of complex networks. Rev. Mod. Phys. 74, 47–97 (2002).
Lieberman, E., Hauert, C. & Nowak, M. A. Evolutionary dynamics on graphs. Nature 433, 312–316 (2005).
Ohtsuki, H., Hauert, C., Lieberman, E. & Nowak, M. A. A simple rule for the evolution of cooperation on graphs and social networks. Nature 441, 502–505 (2006).
Gonzalez, M. C., Hidalgo, C. A. & Barabasi, A.-L. Understanding individual human mobility patterns. Nature 453, 779–782 (2008).
Brockmann, D., Hufnagel, L. & Geisel, T. The scaling laws of human travel. Nature 439, 462–465 (2006).
Skyrms, B. & Pemantle, R. A dynamic model of social network formation. Proc. Natl. Acad. Sci. USA 97, 9340–9346 (2000).
Jackson, M. O. & Watts, A. On the formation of interaction networks in social coordination games. Games Econ. Behav. 41, 265–291 (2002).
Newman, M. E. J. The structure of scientific collaboration networks. Proc. Natl. Acad. Sci. USA 98, 404–409 (2001).
Nowak, M. A., Michor, F. & Iwasa, Y. The linear process of somatic evolution. Proc. Natl. Acad. Sci. USA 100, 14966–14969 (2003).
Barabási, A.-L. Taming complexity. Nat. Phys. 1, 68–70 (2005).
Perc, M. & Szolnoki, A. Coevolutionary games - a mini review. BioSystems 99, 109–125 (2010).
Li, A., Wu, T., Cong, R. & Wang, L. One step memory of group reputation is optimal to promote cooperation in public goods games. Europhys. Lett. 103, 30007 (2013).
Tarnita, C. E., Antal, T., Ohtsuki, H. & Nowak, M. A. Evolutionary dynamics in set structured populations. Proc. Natl. Acad. Sci. USA 106, 8601–8604 (2009).
Antal, T., Ohtsuki, H., Wakeley, J., Taylor, P. D. & Nowak, M. A. Evolution of cooperation by phenotypic similarity. Proc. Natl. Acad. Sci. USA 106, 8597–8600 (2009).
Nowak, M. A., Tarnita, C. E. & Antal, T. Evolutionary dynamics in structured populations. Philos. T. R. Soc. B 365, 19–30 (2010).
Wu, B. et al. Evolution of cooperation on stochastic dynamical networks. PLoS ONE 5, e11187 (2010).
Wu, B., Zhou, D. & Wang, L. Evolutionary dynamics on stochastic evolving networks for multiple-strategy games. Phys. Rev. E 84, 046111 (2011).
Rapoport, A. & Chammah, A. Prisoner's Dilemma: A Study in Conflict and Cooperation. Ann Arbor paperbacks (University of Michigan Press, Ann Arbor, 1965).
Hardin, G. The tragedy of the commons. Science 162, 1243–1248 (1968).
Broom, M., Cannings, C. & Vickers, G. Multi-player matrix games. B. Math. Biol. 59, 931–952 (1997).
Hauert, C., Monte, S., Hofbauer, J. & Sigmund, K. Replicator dynamics for optional public good games. J. Theor. Bio. 218, 187–194 (2002).
Zheng, D. F., Yin, H. P., Chan, C. H. & Hui, P. M. Cooperative behavior in a model of evolutionary snowdrift games with n-person interactions. Europhys. Lett. 80, 18002 (2007).
Souza, M. O., Pacheco, J. M. & Santos, F. C. Evolution of cooperation under n-person snowdrift games. J. Theor. Bio. 260, 581–588 (2009).
Wang, J., Fu, F., Wu, T. & Wang, L. Emergence of social cooperation in threshold public goods games with collective risk. Phys. Rev. E 80, 016101 (2009).
Gokhale, C. & Traulsen, A. Evolutionary games in the multiverse. Proc. Natl. Acad. Sci. USA 107, 5500–5504 (2010).
Pacheco, J. M., Santos, F. C., Souza, M. O. & Skyrms, B. Evolutionary dynamics of collective action in n-person stag hunt dilemmas. Proc. R. Soc. B 276, 315–321 (2009).
Perc, M., Gómez-Gardeñes, J., Szolnoki, A., Floría, L. M. & Moreno, Y. Evolutionary dynamics of group interactions on structured populations: a review. J. R. Soc. Interface 10 (2013).
Wu, B., Traulsen, A. & Gokhale, C. S. Dynamic properties of evolutionary multi-player games in finite populations. Games 4, 182–199 (2013).
Wu, T., Fu, F., Zhang, Y. & Wang, L. The increased risk of joint venture promotes social cooperation. PLoS ONE 8, e63801 (2013).
van Veelen, M. & Nowak, M. A. Multi-player games on the cycle. J. Theor. Bio. 292, 116–128 (2012).
Kurokawa, S. & Ihara, Y. Emergence of cooperation in public goods games. Proc. R. Soc. B 276, 1379–1384 (2009).
Hauert, C., Michor, F., Nowak, M. A. & Doebeli, M. Synergy and discounting of cooperation in social dilemmas. J. Theor. Bio. 239, 195–202 (2006).
Frank, S. A. A general model of the public goods dilemma. J. Evolution. Biol. 23, 1245–1250 (2010).
Archetti, M. & Scheuring, I. Review: Game theory of public goods in one-shot social dilemmas without assortment. J. Theor. Bio. 299, 9–20 (2012).
MacLean, R. C., Fuentes-Hernandez, A., Greig, D., Hurst, L. D. & Gudelj, I. A mixture of “cheats” and “co-operators” can enable maximal group benefit. PLoS Biol. 8, e1000486 (2010).
Hammes, G. G. Enzyme Catalysis and Regulation (Academic Press, New York, USA, 1982).
Santos, F. C., Santos, M. D. & Pacheco, J. M. Social diversity promotes the emergence of cooperation in public goods games. Nature 454, 213–216 (2008).
Wu, B., Altrock, P., Wang, L. & Traulsen, A. Universality of weak selection. Phys. Rev. E 82, 046106 (2010).
Mertzios, G. B. & Spirakis, P. G. Strong bounds for evolution in undirected graphs. CoRR, abs/1211.2384 (2012).
Hauert, C. & Doebeli, M. Spatial structure often inhibits the evolution of cooperation in the snowdrift game. Nature 428, 643–646 (2004).
Taylor, C. & Nowak, M. A. Evolutionary game dynamics with non-uniform interaction rates. Theor. Popul. Biol. 69, 243–252 (2006).
Hamilton, W. D. Innate Social Aptitudes of Man: An Approach from Evolutionary Genetics 133–153 (1975).
Szabó, G. & Fáth, G. Evolutionary games on graphs. Phys. Rep. 446, 97–216 (2007).
Pacheco, J. M., Traulsen, A. & Nowak, M. A. Coevolution of strategy and structure in complex networks with dynamical linking. Phys. Rev. Lett. 97, 258103 (2006).
Allen, B. & Nowak, M. A. Games on graphs. EMS Surv. Math. Sci. 1, 113–151 (2014).
van Veelen, M., García, J., Rand, D. G. & Nowak, M. A. Direct reciprocity in structured populations. Proc. Natl. Acad. Sci. USA 109, 9929–9934 (2012).
Fudenberg, D. & Imhof, L. A. Imitation processes with small mutations. J. Econ. Theory 131, 251–262 (2006).
Wu, B., Gokhale, C. S., Wang, L. & Traulsen, A. How small are small mutation rates? J. Math. Biol. 64, 803–827 (2012).
Do, A.-L., Boccaletti, S. & Gross, T. Graphical notation reveals topological stability criteria for collective dynamics in complex networks. Phys. Rev. Lett. 108, 194102 (2012).
Wu, B., García, J., Hauert, C. & Traulsen, A. Extrapolating weak selection in evolutionary games. PLoS Comput Biol 9, e1003381 (2013).
Gore, J., Youk, H. & van Oudenaarden, A. Snowdrift game dynamics and facultative cheating in yeast. Nature 459, 253–256 (2009).
Acknowledgements
We are indebted to Yong Wang, Guangming Xie, Maozheng Guo, Xi Weng and Te Wu for useful discussions. Comments from Arne Traulsen and the two anonymous referees are acknowledged. We thank James Price to help us improve the clarity and readability of the manuscript. A.L. and L.W. are supported by NSFC (Grants No. 61375120 and No. 61020106005). B.W. acknowledges the sponsorship by the Max-Planck Society.
Author information
Authors and Affiliations
Contributions
A.L., B.W. and L.W. devised and analysed the model. A.L. performed theoretical derivation and numerical simulations. A.L., B.W. and L.W. analysed the results and wrote the paper.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary Information for "Cooperation with both synergistic and local interactions can be worse than each alone"
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Li, A., Wu, B. & Wang, L. Cooperation with both synergistic and local interactions can be worse than each alone. Sci Rep 4, 5536 (2014). https://doi.org/10.1038/srep05536
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep05536
This article is cited by
-
Evolutionary Game Dynamics in a Finite Continental Island Population Model and Emergence of Cooperation
Dynamic Games and Applications (2022)
-
Cooperation in spatial public good games depends on the locality effects of game, adaptation, and punishment
Scientific Reports (2021)
-
Aspiration Can Promote Cooperation in Well-Mixed Populations As in Regular Graphs
Dynamic Games and Applications (2021)
-
Evolution of cooperation on temporal networks
Nature Communications (2020)
-
Chris Cannings: A Life in Games
Dynamic Games and Applications (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
