Abstract
Nature has adapted different methods for surviving dry, arid, xeric conditions. It is the focus of this comparative review to pull together the relevant information gleaned from the literature that could be utilized to design moisture harvesting devices informed by biomimetics. Most water harvesting devices in current use are not informed by nature and those that do are usually based on a biomimetic principle that has been based on one species only. This review draws on the published literature to establish a list of species (animals (vertebrates/invertebrates) and plants) whose habitat is in mainly dry or arid regions and that are known to harvest airborne moisture. Key findings have been outlined and review comments and discussion set out. Following this, surface feature convergences have been identified, namely hexagonal microstructures, groove-like and cone-like geometries. This has been coupled with direction of water flow that is driven by surface energy. As far as the authors are aware, this convergent evolution has not been brought together in this manner before. In the future this information could be translated into an engineered device for collecting water from airborne sources.
Export citation and abstract BibTeX RIS
1. Introduction and thermodynamic background
To exist in xeric conditions, nature has adapted different methods of survival with one of the first known studies of plants absorbing airborne moisture (namely dew and rain) being carried out by Hales (1727). Over a 100 years ago, Lloyd (1905) held the view that it would be future research that would 'throw light' on how plants are able to absorb airborne moisture and, in particular, how the structures and surface properties involved would facilitate this. In support of this, Bolwig (1958) stated that: 'the most important source of water in the Kalahari is from green leaves'. Invertebrates, such as the Namib Desert darkling beetles, make use of fog that rolls in across parts of the Namib Desert from the Atlantic Ocean and it is this form of water survival that is the focus of this review along with dew condensation. Noting the central importance of airborne water, it is first of all important to gain an understanding of the thermodynamics of the psychrometric mechanisms that are present and this will be set out in the following paragraphs.
Rain, fog, mist and dew are all sources of airborne moisture and all are important in the field of water harvesting. Fog and dew both emanate from water vapour in the air condensing to form water droplets, either suspended in air, or formed onto a surface. The size of water droplets also varies with rain droplets ranging from 0.5 to 5 mm and fog droplets between 1 to 40 µm (Fessehaye et al 2014) with a visibility of less than 1 km being the standard for the presence of fog (Met Office 2014).
Fog is visible and has been classified into seven different types, four of these are distinguished by their location and formation process (radiation, sea, steam and advection) and the other three named from their geographical location (coastal, valley and orographic) (Fessehaye et al 2014). The formation of fog usually occurs from local water sources. It originates as vapour from either evaporation from such local sources of water or via the process of transpiration of water from plants. This atmospheric vapour nucleates on condensation nuclei present in the atmosphere such as fine particles of dust and salt, condensing and forming fog droplets. The size of these visible water droplets will vary according to the diameter of the condensation nuclei.
For water to condense onto a surface to form dew, the surface temperature needs to be cooler than or the same as the temperature of the surrounding air. This saturation of water vapour occurs at or below the dew-point temperature and is dependent upon the amount of water vapour that is present in the atmosphere.
The condensing surface temperature is influenced by atmospheric conditions. For example, the night sky acts like a black body having a radiation sink temperature of 2.9 K (Kittel and Kroemer 1980, p 97). This facilitates the formation of dew on a still, clear and cloudless night, on certain surfaces that radiate to this reference temperature through long wave infrared radiation. Nucleation of dew droplets forms under thermodynamically stable conditions which are not evaporating and where the temperature of the substrate is found to be lower than that of the surrounding atmosphere (Beysens 1995).
A drop resting on a solid forms a contact angle (θ) at the three-phase boundary. It is this angle measurement that characterizes surface wettability (Koch and Barthlott 2009) and is illustrated in figure 1.
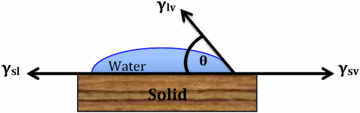
Figure 1. Surface wetting at the solid–liquid–air interface.
Download figure:
Standard image High-resolution imageReferring to figure 1, γlv is the surface tension between the liquid and the gas (or vapour), γsl is the surface tension between the solid and the liquid and γsv is the surface tension between the solid and the gas. Water has a relatively high surface tension (γlv = 72.1 mN m−1). This means that the contact angle for water is higher when compared with a liquid having a lower surface tension. Young's equation (equation (1)) captures this relationship between the contact angle (θ) and the surface tension components (γ)
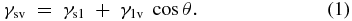
When the contact angle θ is less than 90° surface wetting occurs and the surface is deemed hydrophilic in nature, as it attracts water that also clings to the surface. In contrast, when the liquid molecules become more attracted to each other (due to the cohesive or intermolecular forces pulling the water molecules tighter), wetting of the surface decreases. This is reflected in a wetting angle that exceeds 90° and the surface is considered to be hydrophobic and the droplets become mobile on the surface. Hydrophilic and hydrophobic materials are illustrated in figure 2 in terms of their interaction with a static water droplet.
Figure 2. Hydrophilic and hydrophobic contact angles.
Download figure:
Standard image High-resolution imageIt has been found that the condensing substrate surface properties play an important role in the nucleation process, especially the wettability of the surface (Zhao and Beysens 1995, Andrieu et al 2002).
The surfaces of plants and animals tend to be textured rather than perfectly smooth thus being more heterogeneous in nature. This results in contact angle hysteresis, which is the difference between the advancing contact angle and the receding contact angle (De Gennes et al 2004). The advancing contact angle is the maximum angle an expanding droplet can maintain with stability compared with the receding contact angle, being the minimum angle a retracting droplet can maintain in a stable state. This hysteresis explains why a liquid clings to a surface (Koch and Barthlott 2009). Thus a droplet can be effectively 'pinned to' or 'roll-down' an inclined surface depending on the balance between surface tension and gravitational forces (Quéré 2003). Mention should also be made of the Gibbs' criterion with regard to pinning of water droplets. Gibbs found that at a substrate's sharp edges (such as those at the apex of a cone) Young's equation (equation (1)) no longer held true with a contact angle (α) ranging from θ ≤ α ≤ θ + φ with the substrate's apex edge angle being π – φ (Dutka et al 2012).
For droplets resting on textured surfaces, they can be described as either being in a completely wet Wenzel state (Wenzel 1936) (i.e. whereby the droplet interacts between and on the surface asperities) or a Cassie–Baxter state (Cassie and Baxter 1944) (i.e. where the droplet sits on the top of the surface asperities and thus partially on air that is between the asperities) (Marmur 2003). The adhesive properties of a droplet in the Wenzel state are found to be such that the droplet is 'highly pinned' (Lafuma and Quere 2003).
2. Moisture harvesting in animals and plants
In 1957, a review of the literature was carried out by Stone (1957) looking at dew and the absorption of water in plants in any climate. This review updates and extends the work by Stone to include animals and plants that live in dry and arid climates, where precipitation is severely lacking and where direct dew/fog harvesting is potentially essential to their survival. Several species have been noted to obtain their water source from rain harvesting. The snake Crotalus cerastes laterorepens (Rorabaugh 2007), that can be found in the Yuma Desert, and the tortoise Psammobates tentorius trimeni (Auffenberg 1963), found in semi-arid regions in Southern Africa, both harvest rainwater. However, rainwater harvesting will not be a point of consideration for this review. The current focus is primarily on nature harvesting airborne water (i.e. dew and fog) in regions devoid of rainwater. The first task undertaken was to identify any animals or plants cited as being direct dew or fog harvesters in nature. This information has been collated into tables 1 and 2 along with the relevant citations that report the species' known dew or fog harvesting capability.
Table 1. Animals living in dewy/foggy and arid climes that are cited as harvesting moisture directly.
| Animal | Notes | |
|---|---|---|
| Beetles | Physosterna Cribripesa | Fog basking Namib Desert beetle: 'fog-collecting structure' on beetle's back collects fog water droplets (Parker and Lawrence (2001) renamed to P. cribripes at a later date); Nørgaard and Dacke (2010): questionable fog basker. |
| Onymacris unguicularis | Fog basking Namib Desert beetle (Nørgaard et al 2012, Van Damme 1991): 'assumes the head standing fog basking behaviour' (Nørgaard and Dacke 2010); Hamilton and Seely (1976) observed fog precipitating on the beetle's body; Seely (1979) stated that this beetle species fog basks with a head-down stance'. | |
| Stenocara gracilipes | Namib Desert beetle (Nørgaard and Dacke 2010): debatable fog basker. | |
| Onymacris laevicepsb | Namib Desert beetle that collects fog indirectly from fog dampened surfaces (Nørgaard and Dacke 2010). Drinks water from condensed fog water on vegetation (Seely 1979). | |
| Onymacris bicolor | Namib Desert beetle. 'Well known to fog bask' (Nørgaard and Dacke 2010); Seely (1979) highlighted this beetle's dorsum as the area from which fog water is obtained. | |
| Frog | Litoria caerulea | Tropical northern Australia green tree frog (harvest fog in the dry season). Tracy et al (2011): water condenses onto the frog's skin in a warm hollow (i.e. onto the frog's cooler skin). |
| Lizards | Moloch horridus | Australian thorny devil (agamid) from the Australian deserts. Comanns et al (2011) discuss the lizard's moisture harvesting ability from 'air humidity, fog, dew, rain or even from humid soil'. Gans et al (1982) suggest that condensed dew water on the lizard's skin is their main source of water. |
| Phrynocephalus arabicus | Arabian toadhead agama (agamid) from the arid regions of the Near East and Arabian Peninsula. Comanns et al (2011) discuss the lizard's moisture harvesting ability from 'air humidity, fog, dew, rain or even from humid soil'. | |
| Phrynosoma cornutum | Texas horned lizard (iguanid) from the North American deserts. Comanns et al (2011) discuss the lizard's moisture harvesting ability from 'air humidity, fog, dew, rain or even from humid soil'. | |
| Spider | Uloborus walckenaerius | Cribellate spider's web is able to collect water from humid air (Zheng et al 2010) state that only the wetted silk web is functional for this but others dispute this theory) |
aStenocara gracilipes by some authors. bEven though the beetle Onymacris laeviceps does not collect fog directly, it has been included in this table as Nørgaard and Dacke (2010) studied its elytra for fog water collection efficiency as a comparative study with three other beetle species.
Table 2. Plants living in dewy/foggy and arid climes that are cited as harvesting moisture directly.
| Plant | Notes | |
|---|---|---|
| Trees | Sequoia sempervirens | These Redwood trees from California intercept fog and use this water they harvest, which also adds to the hydrological input of the region (Dawson 1998). |
| Pseudotsuga menziesii | Douglas fir tree: Ingwersen (1985) (cited in Dawson 1998) discussed how the amount of water from the fog decreased in the ecosystem if trees (like Douglas fir (Pseudotsuga menziesii)) were removed from the watershed. | |
| Namib plants | Welwitschia mirabilis | Namib Desert plant: Van Damme (1991) comments that the Welwitschia plant may be a fog harvester but this is debatable. |
| Trianthema hereroensis (Aizooceae) | This plant only grows in the Namib Desert in the area where fog is present. Two papers site their use of fog: 'Rapidly absorbs fog water through its leaves' (Van Damme 1991); 'uses fog water absorbed through its leaves (Seely 1979). | |
| Stipagrostis sabulicola | Namib dune bushman grass (Poaceae). Fog and possible dew harvester. Ebner et al (2011) report that S. sabulicola 'is able to harvest substantial amounts of water from nocturnal fog events' with water droplets forming on the leaves; Nørgaard et al (2012) state that the grass collects water directly from fog; obtains moisture through its root system (Seely 1979, Van Damme 1991). Roth-Nebelsick et al (2012) mention the fog collecting structure of this plant. | |
| Cacti | Opuntia microdasys | Chihuahua Desert cactus. Has an 'efficient fog collection system' (Ju et al 2012). |
| Copiapoa haseltoniana | This cacti uses dew that condenses on its spines (Nobel 2003). | |
| Discocactus horstii | These three cacti are known for their uptake of water from air by their spines (Schill and Barthlott 1973, Schill et al 1972, Barthlott and Capesius 1974, all cited in Nobel (2003)). | |
| Turbinicarpus schmiedickeanus klinkerianus | ||
| Mammillaria theresae | ||
| Eulychnias | These genera (in which there are five species) of cacti from Chile are known to use dew and fog water to survive (Yetman 2007). | |
Van Damme (1991) states that the Namib Desert shrub known as the Arthraerua leubnitziae (Amaranthaceae) may use fog water indirectly (but the paper does not discuss in what way exactly). Due to this species being an indirect harvester, it will not be considered relevant for this review.
3. Overview of key findings from the literature
A further review of the literature was undertaken to source research on any of the named species in tables 1 and 2, with regard to dew/fog/moisture harvesting. This section summarizes the key findings from the identified papers, with the focus only on direct fog or dew harvesting species. For ease of reading the findings considered relevant have been split into five subsections: beetles, frogs, lizards, spiders and plants. In performing this review, the format adopted in the following paragraphs is identifying each paper (i.e. the title and author/s), typically followed by a few sentences that summarize the paper together with their claims under 'key findings' and this paper's discussion under 'review comments' where appropriate. Some papers are based on observations and others on volumetric means.
3.1. Beetles
Five beetles have been identified as harvesting dew or fog, four directly (Physosterna cribripes, Onymacris unguicularis, Stenocara gracilipes and Onymacris bicolor) and one indirectly (Onymacris laeviceps which has been included here as Nørgaard et al (2012) studied its fog harvesting efficiency in comparison with three of the direct water harvesters).
There seems to be general agreement with regard to the two Onymacris beetle species being direct fog harvesters, but there is some debate in the literature whether or not Phyosterna cribripes and Stenocara gracilipes harvest fog directly. Parker and Lawrence (2001) discuss the fog-basking stance of Physosterna cribripes' (renamed at a later date from Stenocara sp.) which has alternatively been suggested as possibly being the beetle's alarm call behaviour (Nørgaard and Dacke 2010). However, following the 2001 Parker and Lawrence article, much has been gained in the study and surface replication of the Physosterna cribripes' bumpy, hydrophilic and hydrophobic nature (Zhai et al 2006, Garrod et al 2007, Dorrer and Ruhe 2008, Hong et al 2012) which has further informed the field of moisture harvesting.
Both Physosterna cribripes and Stenocara gracilipes have black elytras (i.e. the beetle's hard exoskeleton covering the dorsal surface of their abdomen, 'wing covers') with a covering of bumps with slopes and valleys in between, but Stenocara gracilipes has a smaller abdomen and thus a smaller surface area. Onymacris unguicularis has a black elytra with grooves that run over only part of the elytra (i.e. from back to middle). Similarly, Onymacris laeviceps has grooves that are finer than those of Onymacris unguicularis and that run from the back to the front of the elytra. Onymacris bicolor is the only beetle that has white elytra. The largest of the five beetles is Physosterna cribripes and the smallest is Stenocara gracilipes. The literature reviewed only focused on four of the five beetles (no papers were found examining the moisture harvesting of Onymacris bicolor in any more detail than that given in table 1).
Two of the papers reviewed in this section discuss the hydrophobic and hydrophilic properties of Physosterna cribripes. Parker and Lawrence (2001) found the bumps to be hydrophilic, whereas Nørgaard et al (2012) stated that there were no hydrophilic areas anywhere on any of the beetle species (Physosterna cribripes, Onymacris unguicularis, Stenocara gracilipes or Onymacris laeviceps). If Nørgaard et al (2012) are right, the condensing water should not show any preferential nucleation, whereas if the elytra surface is hydrophilic and hydrophobic in nature, then preferential nucleation should be observed on the hydrophilic bumps as was observed on the synthetic surfaces utilized in a study by Varanasi et al (2009). However, Guadarrama-Cetina et al (personal communication) found the valleys to be the primary nucleation sites which they concluded to be due to a hexagonal microstructure, rather than to the surface having a hydrophilic nature.
From the literature, no general consensus seems to have been reached with regard to the effect of wax on the beetle's elytra which would indicate hydrophobic areas. However, the main focus has been on just one species of beetle, namely Physosterna cribripes. Furthermore, work has been conducted using dead beetles, live beetle elytra may have different surface properties that will have an impact on flow in moisture harvesting. A possible resolution could be to examine the elytra surface properties to gain an understanding of areas that are hydrophilic and this will be discussed later.
3.1.1. Fog-basking behaviour and water collection efficiency in Namib Desert darkling beetles (Nørgaard and Dacke 2010)
The behaviour (from live specimens) of four different genera of Namib Desert beetles (Physosterna cribripes, Onymacris unguicularis, Stenocara gracilipes, Onymacris laeviceps) was noted through observation.
Key findings. Of the live beetles, only Onymacris unguicularis exhibited the classic fog basking behaviour, triggered by the presence of fog. Also the elytra of dead specimens were analysed for water collection efficiency, achieved by volumetric means using an Eppendorf tube to collect fog water run-off. No significant difference was found in the quantity of fog water collected from the elytra of all four species of beetle, even though each possesses different surface structures and areas. After the study accounted for the different beetle sizes by normalising the data to the surface area of Physosterna cribripes (as it is the largest of the four), it was found that the largest beetle collected the least water; Onymacris unguicularis and Stenocara gracilipes were the most efficient fog harvesters. Stenocara gracilipes, the smallest of the four beetles, harvested the most water.
It was found that Stenocara gracilipes harvested nearly twice the amount of water per surface area unit when compared with the larger Physosterna Cribripes. These two beetles, possessing bumpy elytras, came out at both ends of the water collecting efficiency spectrum, compared to the other two species of beetles having grooved elytras. Dead specimens of all four beetles were found to have no areas on their elytra that were hydrophilic (determined using the Sudan III staining method to identify waxy areas). Only Physosterna cribripes exhibited hydrophobic waxy peaks.
The authors suggested that the smaller beetles have a thinner boundary layer and as such make better fog harvesters, in a similar way to small leaves that harvest fog. Boundary layer growth due to air flow is indicated by a Reynolds number for which the beetle size is the characteristic dimension (Bonan 2002).
Review comments. The surface area normalization does not take into account the microstructure of the beetles' elytron. Observing the photos taken from the SEM, it can be seen that Stenocara gracilipes (the most efficient fog harvester) has more foci (i.e. surface asperities), that is, there appear to be more bumps on the elytra, small and large, when compared to the other three beetle species.
With regard to the identified waxy elytra areas, wax can be seen from the Sudan III staining method on the bumps of the Physosterna cribripes showing them to be hydrophobic but if, as the authors stated, 'no hydrophilic areas could be identified' on any of the four beetles under examination, then the Sudan III staining method should show wax all over the elytra.
However, examining the figure in the paper, it can be seen that waxy areas are shiny and that there are other areas that are matt and wax free, suggesting hydrophilic areas (see figure 3).

Figure 3. Physosterna cribripes waxy areas. (Reproduced with permission from Nørgaard and Dacke 2010: figure 3, p 5)
Download figure:
Standard image High-resolution imageTwo different staining dyes were used to ascertain the waxy areas. Nørgaard et al (2012) carried out the Sudan III method and Parker and Lawrence (2001) utilized the Red O staining method. Red O produces a darker more prominent stain colour of red compared to a lighter orange–red stain colour for Sudan III (Guigui and Beaudoin 2007) thus making Red O more popular (Sigma 2014).
Parker and Lawrence (2001) mention that videos were taken showing water droplets growing on the bumps before rolling down the elytra; the nucleation of water would suggest that this area of the beetles' elytron is hydrophilic. It could also be possible that, if examined under a higher magnification, water condenses first in the valleys and then uses the hydrophobic bumps to guide and roll the condensate down the elytra to the beetle's mouth. The hexagonally arranged structure within the valleys, observed using the SEM, could also indicate hydrophilic areas attributed to topography (see section 4.1 for further discussion and images).
3.1.2. Animal or plant: which is the better fog water collector? (Nørgaard et al 2012)
This study was a volumetric comparison between a beetle (Onymacris unguicularis) and dune bushman grass (Stipagrostris sabulicola) to determine the more efficient fog collector. Ideally the Stipagrostris sabulicola would be discussed in the plant section below but, due to this review paper considering both the beetle and the plant, the key findings are outlined below.
Key findings. The beetles collect fog when it is located at the top of dunes, in contrast to the grass, which collects fog from the base of dunes even though the availability of fog decreases below the ridge. The water run-off dynamics of the beetles was at a steady rate compared to episodic, cascading run off for the grass. The fog water collected from the beetle was much less than that of the grass but interestingly the authors found that a metal wire collected the same amount of water as that of the grass. They stated:
This suggest(s) that the three dimensional structure of the grass straw—rather than any particular surface properties—is the important factor for its water collecting abilities. (p 3).
Initially the beetles collected more water but, after 100 min, the grass suddenly increased its water run-off rate. The authors considered that the stochastic water collecting rate for the grass compared to the beetles' steady rate raises questions about the mechanisms that cause this step change in harvesting capability.
Review comments. The authors conclude that the grass is a significantly better water collector but, this is only true after the 100 min mark when the grass cascades water, increasing its water collection fivefold in the subsequent 20 min. It could be argued that, under natural conditions, the beetles are only able to drink a limited amount of water and at a certain constant rate, after which they would have reached their holding capacity (the subelytra gap below the beetle's elytra and dorsum area enables the beetle to drink and expand (Slobodchikoff and Wismann 1981) but possibly only until this chamber area is full). They would also have to be efficient at collecting water during that time period to minimize being in an exposed place for an extended period and consequently vulnerable to predators. The grass on the other hand is sessile and as such its water requirements are different.
A run off rate that changes so dramatically after 100 min appears out of place with the rest of the experiment up to this time-marker and is unexplained by the authors other than stating that the grass cannot move in the same way as the beetle. The question arises: is this due to an experimental system error or is it due to the grass becoming saturated? The fact that the authors repeated the experiment with twelve grass straws would rule out experimental system error. The fact that the 100 mm grass sections were no longer attached to their root system could have impacted on their innate properties, somehow creating sudden run-off rate.
As no SEM images of the grass structure were provided, no comparison could be made at the micron level, which would have been useful in understanding the surface architecture and comparing the results with other literature. The researchers did, however, compare the specimens under examination by taking into account the upper surface area, which ensured a fair comparison.
With regard to the metal wire (i.e. galvanized iron) being found to harvest the same amount of fog water as the grass specimen, this could be due to the temperature of the metal causing dew water to be harvested, if the metal was cold enough. Material should be considered a factor. A picture of the metal wire in question was not given by the authors so it is difficult to comment on whether its fog collecting ability could be due in part to surface roughness and consequently more foci (surface asperities) to act as points for condensation. It would have been interesting to view SEM images of the wire to be able to comment further and draw further analyses from the results.
3.1.3. Water capture by a desert beetle (Parker and Lawrence 2001)
Even though this paper classified the beetle under examination as Stenocara, the authors later reclassified it as Physosterna cribripes. They examined the water collecting ability of this beetle in foggy conditions.
Key findings. The surface structure is bumpy with alternating regions of hydrophilic areas (smooth non-waxy areas seen as bumps) and hydrophobic (textured waxy regions seen as valleys and the slopes of the bumps'). The textured waxy regions of the beetle's elytra are super-hydrophobic in nature, displaying a hexagonal array of flattened hemispheres at a micro level (Parker and Lawrence 2001, figure 1(c), p 33).
By capturing video footage, it was observed that a water droplet forms quickly on the hydrophilic peak until it covers the entire waxy area upon which the water droplet rolls off the peak and down the angled elytra (due to the beetle's fog-harvesting stance). It is 'guided by the slight purchase afforded by other peaks along its path'. The rolling drop must reach a particular radius to overcome the force of the oncoming fog bearing wind (recorded as 5 m s−1 in the Namib Desert) in its natural desert environment.
The authors examined the beetle structure further by replicating it crudely on a microscope slide using glass spheres half embedded in wax. They tested four different slide structure combinations against each other for wind-blown mist water collecting efficiency. They found the square array of beads to be significantly the best collector and the plain glass slide the most variable. This led them to the conclusion that having a combination of hydrophilic and hydrophobic points on a surface yielded the best water collection from mist.
Review comments. The researchers were the first to look closely at the Namib beetle as holding potential clues to aid in fog harvesting for humans. Even though the beetle species was misnamed in the article and corrected at a later date, the authors included a full picture of the species under investigation, which enabled subsequent researchers to identify the specimen correctly. Future research papers on species of any kind should, as good practice, include a full picture in this way to ensure that species are identified correctly.
Having studied the water collection by exposing samples to a fine water spray (this spray was not characterized in terms of droplet diameter that has a typical diameter of 1–40 µm for fog) a further step could have been to look at samples in a fog chamber, giving even more insight through better representation of climatic conditions. The researchers concluded that the square array structure on the microscope slides yielded the best results but the plain glass slide yielded the same average quantity of water so long as the water ran straight into the collection vial. It might be a useful consideration when designing a fog harvester or carrying out such experiments, that guttering is used (as is found in the tortoises discussed by Auffenberg (1963)) as a critical element to ensure no captured water is lost.
3.1.4. Dew collection by the Namibian beetle Physasterna cribripes (Tenebrionidae) (Guadarrama-Cetina et al (personal communication))
The ability of the black bumpy elytra of a preserved Physosterna cribripes was examined for its ability to harvest dew water using a condensation chamber to encourage nucleation of water droplets on the elytra, together with a scanning pyrometer to study its IR emissivity.
Key findings. Condensation was always observed to start in the hydrophilic 'valleys' of the elytra with water condensing on the hydrophobic 'bumps' later. It was postulated that this was due to:
- (i)'thermal effects' with cooler valley regions
- (ii)the surface roughness and wettability creating a difference in the nucleation barrier between the valleys and bumps.
The surface properties of the elytra were concluded to be the cause of dew formation, namely the hexagonal microstructure along with its high IR emissivity (0.95 ± 0.07 between the wavelength domain 8–14 µm). The wettability difference between the bumps and valleys was obtained from the contact angle data, with the contact angle being higher for the bumps (i.e. 124° ± 4°) compared with the valleys (i.e. 64° ± 8°) reflecting their hydrophobic and hydrophilic behaviour respectively.
Review comments. The authors could not explain why the valleys appeared 'more hydrophilic than the bumps' as wax was found to coat both areas. They attributed the wettability to be due to the microstructure. It could also be the case that this wax, which is being presumed to be hydrophobic, is in fact hydrophilic in nature, as is found to be the case for plant waxes (Wagner et al 2003) cited in Roth-Nebelsick et al (2012)). A waxy coating found on the surface of animals in xeric conditions is known to minimize desiccation (Guadarrama-Cetina et al (personal communication)), the same is true for plants. It could thus be said that if these surface waxes were found to be hydrophilic in nature for the beetles under investigation (as is already known for wax on the plants surface), this could encourage the nucleation of water droplets that in turn would coalesce and coat the surfaces of the plants and animals. This would also act as a barrier to evaporation (for animals) and transpiration (for plants), effectively minimizing water loss.
3.2. Frogs
Only one frog was identified from the literature as being noteworthy for this review, namely the Litoria caerulea, an Australian green tree frog
3.2.1. Natural history note: condensation onto the skin as a means for water gain by tree frogs in tropical Australia (Tracy et al 2011)
Green tree frogs (Litoria caerulea) from the Northern Australian tropics were studied for water condensation onto their skin in warm natural and artificial tree hollows.
Key findings. The frog's body mass increased consistently (due to water gain) when moving from the cool exterior environment to the warm humid natural tree hollow, with condensation being observed on the frog's dorsal skin. Similarly the frog's mass increased when entering an artificial tree hollow, again displaying condensation on the dorsal skin.
As long as conditions allowed (high humidity, radiant heat exchange, cold sky), dew formed on the frog's skin (the temperature of which was below the dew point) before the frog entered the hollow.
Review comments. This study focused only on dew formation and not fog harvesting. It has been included in this review as this Australian species has to survive with little, if any, free water during the extreme conditions of the dry season. It seems to manage this by harvesting dew on its skin and as such was considered an useful inclusion to this study.
The body temperature of the frog seemed to be the critical factor in dew harvesting and the authors quote a number of other references whereby species have been stated as utilizing their body as rain or dew harvesters:
- Lizards: Bentley and Blumer (1962) and Gans et al (1982),
- Snakes: Sherbrooke (2004) and Glaudas (2009)
- Tortoises: Auffenberg (1963).
- Insects: Hamilton and Seely (1976), Hamilton et al (2003) and Henschel and Seely (2008)).
Further analysis of this frog species will not be carried out in this review as it is not seen as central to fog harvesting and the reviewed paper did not give details of the frog's skin microstructure.
3.3. Lizards
Three lizards were identified as fog harvesters (along with being harvesters of moisture in other forms too), namely Moloch horridus, Phrynosoma cornutum and Phrynocephalus arabicus. The lizards originate from different locations worldwide and are deemed to have evolved their moisture harvesting abilities independently (Comanns et al 2011).
3.3.1. Moisture harvesting and water transport through specialized micro-structures on the integument of lizards (Comanns et al 2011)
This research focused on three different types of lizards that harvest water (from air humidity, fog, dew, rain or soil) from arid regions of the globe, namely: Moloch horridus, Phrynocephalus arabicus and Phrynosoma cornutum. They chose these three lizards as they felt that each had evolved their water harvesting abilities independently and so the surface micro-structure of each was compared for convergences.
Key findings. The lizard's skin did not take in water significantly, instead it acted as a transporter of water to the lizard's mouth. The outer beta keratin layer of the lizard's skin possessed a 'honeycomb' structure (being the least pronounced in Phrynocephalus arabicus). This outer surface was found to be superhydrophilic in nature, displaying a highly wettable state, which the authors found to be caused by structure rather than material.
Even though all three lizards had totally different external forms, all possessed scales that were highly wettable, with spreading of water at high velocity. Phrynosoma cornutum illustrated a greater forward water velocity towards the lizard's mouth compared with the other two lizard species.
Early morning water condensation may be the result of these cold-blooded lizards' body temperature remaining low following extreme cold nights. The researchers tested the condensation effect of the scales' honeycomb surface by creating replica epoxy surfaces and found it increased condensation by 100% compared to non-structured epoxy surfaces. They stated:
This might be caused by the increased roughness only, because a rough surface supports more condensation foci. However, if condensation had taken place leading to a water film in the honeycombs, the surface exhibits the above described high wettability which would then allow for further improved moisture harvesting from other water sources like rain, fog or dew. (p 210).
Review comments. It would have been interesting from the perspective of this review if the research group had also carried out a test for wax on the surface of the lizards. The lizard's outer surface is made up from beta keratin, known for its high strength to weight ratio, along with chitin (which constitutes a beetle's elytra), which is similar to keratin. It is interesting to note that a spider's web is also well known for its strength and is also made from keratin.
3.4. Spiders
The Ulborus walckenaerius's wetted silk web has been identified as not only a water collector but a directional one.
3.4.1. Directional water collection on wetted spider silk (Zheng et al 2010)
These researchers observed the formation and collection of water droplets on the Uloborus walckenaerius spider's web. They then replicated this web through the use of a porous synthetic material in combination with an artificial knot periodicity of 394.6 ± 16.1 mm.
Key findings. The silk webs are made from hydrophilic, humidity sensitive proteins that 'enhance the wettability of spider silk, which is favourable for condensing water drops'. Upon wetting, the silk web's structure changes, causing water to be collecteds in a directional manner.
The wetted spider silk displays periodic rough knots with joints in between that display parallel structures. The structure causes directional flow of water, as well as acting as a driving force (due to the surface energy gradient and the Laplace pressure4 ). Water condenses on both the knots and joints but the joint topography acts as a water transporter to the collection sites of the knotted areas. The joints have larger water contact angles when compared with the knotted areas. The knots are more hydrophilic than the joints and they have a higher apparent surface energy. This ties in with Young's equation (equation (1)), for a lower surface energy the contact angle increases thus the hydrophobicity of the material increases (Nakajima et al 2001).
Due to there being an apparent difference in roughness between the knots and joints, water droplets are driven by the surface energy gradient from the lower surface energy, hydrophilic joint to the more hydrophilic knot. The conical knots (i.e. two cones fused together) 'generate a difference in Laplace pressure' thus adding to the directional flow of the condensed water droplets. Spreading of the water droplets occurs more easily on the joints' aligned parallel surface topography compared with the knots, since the joints have a lower hysteresis due to a three phase contact line that is continuous for the joints and discontinuous for the rougher knots.
Review comments. The authors refer to the literature that highlights the 'unusual structural features' and comment that:
Many biological surfaces in both the plant and animal kingdom possess unusual structural features at the micro- and nanometre scale that control their interaction with water and hence wettability. (p 640).
However, this review has compared the surfaces of the species under consideration and highlighted the convergences in section 4 for both plant and animal species.
3.5. Plants
Three plants have been identified as fog harvesters (Opuntia microdasys, Stipagrostis sabulicola and Trianthema hereroensis) and another as a possible fog harvester (Welwitschia mirabilis) together with the Redwood Trees (i.e. Sequioa sempervirens). The Bushman grass (Stipagrostis sabulicola) will not be covered in this section, having already been discussed in the beetle section, as it was part of a comparative study.
3.5.1. A multi-structural and multi-functional integrated fog collection system in cactus (Ju et al 2012)
A single spine from the Chihuahua Desert cactus, Opuntia microdasys, was studied for its fog harvesting abilities and this information is being used by the authors to develop an artificial fog harvesting system.
Key findings. There are three different surface structural features on the cactus spine: the barbed tip, the middle section grooves and the trichomes at the base. Each of these has its own role in fog collection. Droplets of water always moved from the tip to the base regardless of the spine's orientation, thus illustrating the fact that the direction of the water collection on the spine was not affected by gravitational force. The researchers concluded that the water droplets were driven in this direction due to:
- 'The gradient of the surface-free energy' arising from the spine's surface roughness determined by the microgrooves (as there are more grooves near the spine's tip compared to the base, the tip is rougher)
- 'The gradient of the Laplace pressure,' due to the spine's conical shape, enhanced by the spine's roughness
- The asymmetrical surface structure of the barbs causing the 'unidirectional rolling and spreading of the water drops, further adding to the directional movement of the water drops'. (p 3).
Water transportation along the spines was found to be slow compared to the absorption of water by the trichomes. This fast absorption rate enabled more fog water to be harvested quickly.
The spine tip also has a gradient of wettability caused by the gradient of roughness (i.e. due to the grooves). The spine's aligned grooves:
- reduce the water droplet's ability to spread
- enhance the gradient of the Laplace pressure
- generate an anisotropic contact angle hysteresis parallel or perpendicular to the grooves, making the water droplets more likely to move parallel to the aligned grooves and aiding movement of the water towards the spine base.
Multiple spine clusters were not examined but it was postulated that 'the array of multiple clusters on the stem surface may further enhance the fog collection ability'.
Review comments. Another subject not discussed in much detail was the presence of waxy areas on the cactus spine. Again this would have been useful information in order to make a comparison with the other plant and animal species.
3.5.2. Leaf surface structures enable the endemic Namib Desert grass Stipagrostis sabulicola to irrigate itself with fog water (Roth-Nebelsick et al 2012)
The stem or culm of the Stipagrostis sabulicola, has leaves that wrap tightly around it. The structure of the leaf was studied for its fog collecting abilities. This study involved field observations of the plant together with laboratory analysis.
Key findings. The leaf surface has an extremely irregular construction, with a layer of wax, but was found to be hydrophilic with an advancing contact angle that was slightly hydrophobic. Fog water droplets can be seen forming on the tall leaves, and these were found to merge with other droplets that were also clinging to the leaf. Droplets eventually slide down the leaf to the root system upon reaching a critical size, gathering smaller droplets on the way. The water droplet flow is parallel to the grooves on the leaves.
Harvested water droplets were found to run down the surface of the leaf to its base even when the leaf was tilted from its vertical position. This was not the case if the leaf was at an angle of less than 15° to the horizontal. It is suggested that the surface irregularities (i.e. from prickly hairs, waxy structures and possibly deposited sediment such as salt and sand) observed on the leaf could affect the behaviour of hanging droplets. Irregularities could act as points of contact to which small droplets are 'pinned'. This pinning of water droplets aids the formation of larger droplets.
The water collection rate for the leaf was compared with that of nylon bristles of similar dimensions and the leaf proved to be a significantly better collector. It is thought that this is due to the higher surface roughness of the plant compared to the nylon. The plant's structural surface properties are believed to be the reason why water droplets behave in a certain way when interacting with the leaf, with fog water preferring to stick to the grass's furrows first and then coalescing further before reaching a critical mass and rolling down the plant stem.
The authors cite Wagner et al (2003) on the fact that plant waxes possess hydrophilic groups with intrinsic contact angles found to be less than 90°. Gravity, Laplace pressure and surface roughness all play a role in the dynamics of the harvested water droplets.
3.5.3. Fog in the California redwood forest: ecosystem inputs and use by plants (Dawson 1998)
This was a three-year study that looked at the effect of fog water on the ecosystem of the redwood (Sequioa sempervirens) forests of California.
Key findings. Sequioa sempervirens trees were found to intercept fog, which then dripped off the trees, making up 34% of the average annual water input for the forest, with only 17% of the forest average annual water input from fog being recorded when there were no trees present. This paper referred to a paper by Ingwersen (1985) that discussed how the amount of water from fog decreased in the ecosystem through fog drip and stream flow if trees (like the Douglas fir (Pseudotsuga menziesii)) were removed from the watershed.
The study calculated the varying isotopic composition found for fog water compared with other water sources (e.g. rainwater, deep soil water) in this region to work out the quantity of each water source by the redwood forest. It is known that fog tends to have more of the heavier isotopes (such as 2H and 18O) compared to its atmospheric vapour derivative. Furthermore, the authors state that, compared with rainwater, fog is more isotopically enriched due to the fact that the water source of rainwater tends to be different and has depleted levels of H and O because it has travelled a greater distance and because 'water continuously condenses out of the clouds'.
The canopies of tall trees themselves seem to intercept fog, thus enabling fog water to add to the hydrology of the local ecosystem. The authors stated that, when forests are not fully intact, either due to artificial (such as clearance felling), or naturally occurring tree loss, the amount of fog drip declines causing the forest moisture content to decrease along with nutrient content.
Review comments. The Redwood and Douglas fir trees both have cone-like surface features. When examining plants and animals for their ability to harvest fog and dew, examining the isotopic composition of the collected water could prove to be a valuable tool. The isotopic composition would vary for different parts of the world and as such each geographical location would require analysis of the different water sources beforehand. Utilizing a Local Meteoric Water Line (LMWL), as was carried out in this particular study and favoured over the Global Meteoric Water Line (GMWL) to enable a more appropriate isotopic measurement of the local airborne moisture variation, could be a point of consideration for future research in this field. The GMWL is represented in an equation that gives the global average relationship between stable H and O isotopic ratios (Craig 1961).
4. Convergences
With regard to dew and fog harvesting devices, it appears that researchers have until now focused mainly on one species at a time to obtain the secrets of its water harvesting abilities, with the aim of translating this into an engineered device. To date, there has been no really thorough search for convergent mechanisms or microstructures across all species to study dew and fog harvesting. Only the one study by Nørgaard et al (2012) was found which compared an animal with a plant in this regard, but this lacked SEM images and an analysis of structure. This review has looked at the available images of micro-structures of the fog harvesting species under consideration and it appears that these structures seem to be far from unusual. In fact there appear to be considerable parallels across species.
Structure fulfils several functions, including a defence mechanism against predators. This is clearly the case for the spiky lizards and is also a possibility for the cactus. The beetle's hard outer shell also acts as a protective casing that can protect it. It is important that these additional functions are kept in mind whilst examining the fog harvesting abilities of each of these species. A number of the general review papers have mentioned surface structure as an important factor in the water collecting process; several characteristics seem to be arising which are discussed below.
4.1. Groove-like structures
SEM images illustrating groove-like structures were captured by the relevant authors in many instances and were found in the Opuntia microdasys cactus, two species of beetles (Onymacris unguicularis and Onymacris laeviceps) and the lizard Moloch horridus. The Uloborus walckenaerius spider's web was described by the authors as having aligned nanofibril joints which could possibly act in a similar manner to groove-like structures. Examples of some of these structures are seen in figure 4.

Figure 4. Groove-like structures. Different fog harvesting species demonstrate aligned grooves in their structure. Pictures taken by the different authors are shown above. (a) Although not aligned, there are deep groove-like channels in between the scales of the lizard Moloch horridus (reproduced with permission from Gans et al 1982: figure 3(a), p 61 (© 1982 Koninklijke Brill NV)); (b) beetle elytra Onymacris unguicularis and (c) Onymacris laeviceps (reproduced with permission from Nørgaard and Dacke 2010: figures 2 (A2) and (B2), p 4); (d) and (e) the Namib grass Stipagrostis sabulicola leaves possess a grooved surface (reproduced with permission from Roth-Nebelsick et al 2012: figures 1(b) and (c), p 2 (© 2012 The Royal Society)).
Download figure:
Standard image High-resolution imageThese structures are thought to act in all cases to direct water. In the case of the lizard the grooves are not parallel whereas, in the other cases, all the grooves can be seen to be aligned and parallel. The lizard's grooves can be observed to be deeper than those of the other species.
The aligned groove arrangement aids the directional flow of water parallel to the grooves due to an anisotropic contact angle hysteresis as discussed by Ju et al (2012) along with the fact that the 'pinning' of water droplets does not really occur parallel to the grooves and is only really found perpendicular to the grooves (Roth-Nebelsick et al 2012).
These grooves seem to direct the harvested water, thus minimizing water loss (through evaporation) and increasing water capture by moving water away from harvest locations (such as hydrophilic condensation areas) to make room for more water to be harvested. Therefore, the function of the grooves is assumed not to be one of drawing water from the air but rather as a guiding channel for the water drops that have formed previously.
4.2. Cone-like structures
It is immediately apparent that the cactus spine displays a cone-like shape that, according to Ju et al (2012), gives rise to a Laplace pressure gradient which, in turn, is one of the driving forces to direct the flow of the harvested water. Examining the structure of the spider's web, the lizard's scales and the beetle's elytra (Physosterna cribripes and Stenocara gracilipes), it can be seen that they all possess cone-like structures that could act in a similar manner (some of these structures are shown in figure 5 below). In fact, Zheng et al (2010) also highlight that the conical shape acts as a driving force for the collected water.
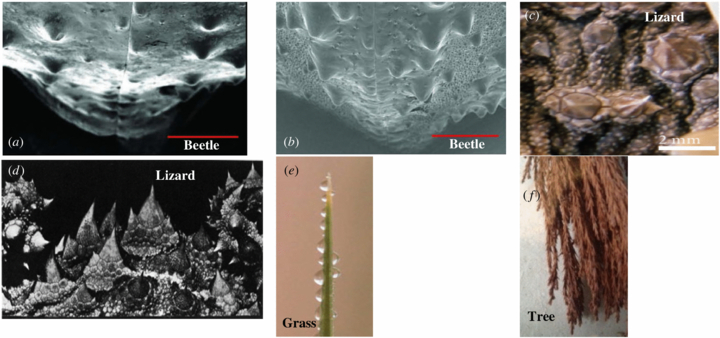
Figure 5. Cone-like structures. Cone structures are evident from the SEM pictures published on different fog harvesting species. (a) The beetle Physosterna cribripes elytra (reproduced with permission from Nørgaard and Dacke 2010: figure 2(D2), p 4); (b) the beetle Stenocara gracilipes, elytra (reproduced with permission from Nørgaard and Dacke 2010: figure 2(C2), p 4); (c) thorny cone-like scales of the lizard Phrynosoma cornutum (reproduced with permission from Comanns et al 2011: figure 2(C), p 206); (d) thorny cone-like scales of the lizard Moloch horridus (reproduced with permission from Gans et al 1982: figure 2(a), p 60 (© 1982 Koninklijke Brill NC)); (e) The pointed tip of the Stipagrostis sabulicola Namib grass (cropped version of Roth-Nebelsick et al 2012: figure 3, p 4 (© 2012 The Royal Society)); (f) Cone-like structures are clearly visible on the leaves of the Sequioa sempervirens (Redwood tree).
Download figure:
Standard image High-resolution imageInterestingly, the water is directed from the tip of the cactus spine to its base regardless of the orientation of the spine. Similarly, a cone shape has been observed in the Uloborus walckenaerius spider's wetted web where water can be observed being directed from the tip (i.e. joint) to the base (i.e. knot).
The Stipagrostis sabulicola grass found in the hyper arid regions of the Namib Desert also clearly exhibits cone-like structures from its tip to its base. This grass similarly directs harvested water from its tip to its base for all angles of the plants leaf greater than 15° to the horizontal. The two species of tree mentioned earlier, the Sequioa sempervirens (Redwood tree) and the Pseudotsuga menziesii (Douglas fir tree) both possess cone-like structures in their needle-like leaf make up.
4.3. Directional water flow
Water drops move in a fixed direction along cone-like structures, as can be seen in figure 6 for the grass and a lizard's skin.

Figure 6. Directed water flow. (a) Fog water droplets form on the Stipagrostis sabulicola grass leaves and run down towards the root system (cropped version of Roth-Nebelsick et al (2012): figure 3, p 4 (© 2012 The Royal Society)); (b) water is directed at high velocity to the mouth of the lizard Phrynosoma (reproduced with permission from Comanns et al 2011: figure 8, Phrynosoma after 3 s, p 211). Note: arrows have been added for clarity.
Download figure:
Standard image High-resolution imageOne of the other points for consideration in addition to the cone-like macro-shape is the roughness gradient of each surface at the micro level, a direct result of the surface asperities. Both Zheng et al (2010) and Ju et al (2012) agree that the surface gradient directs the flow of water for the cactus and the spider's web respectively from a lower surface gradient to a higher surface gradient area. Ju et al (2012) discussed the use of surface energy to drive water from a lower to a higher surface energy. They revealed that the water flows from the more hydrophobic, rougher, less wettable, lower surface energy to the base which has a higher surface energy. Zheng et al (2010) also agreed that the movement of water along a spider's web was towards a higher surface energy, with the knot being more hydrophilic than the joint.
It is known that water can move against gravity due to a surfaces free energy (Chaudhury and Whitesides 1992). This directional flow of water can be put into context using the table 3 below which illustrates the relationship that hydrophilic and hydrophobic surfaces have with regard to the contact angle, surface wettability, water flow and the adhesive and cohesive forces.
Table 3. Overview of wettability and contact angle. FA is the force required for molecules from different surfaces to stick to each other. FC is force required for molecules of the same substance stick to each other.
 |
With regard to the surface roughness, however, the water moves from the rough cactus tip to the smoother base which is in contrast to the spider's web, where water moves from the smoother joint towards the rougher knot area. It should be considered that the nanofibril alignment found on the web's joint area could override the roughness element. However, this water movement from a smaller radius on a cone shape to the larger radius is driven by the Laplace pressure gradient, thus as the curvature of a circle (κ) is the inverse of its radius (κ = 1/r), water will move away from the higher curvature, as was found for drops on a conical wire (Lorenceau and Quéré 2004).
Directed water flow for the grass Stipagrostis sabulicola was achieved mainly by 'relief and surface roughness' (Roth-Nebelsick et al 2012). The grass leaf was found not to possess wettability gradients and as such this was not a factor in the directed flow (Roth-Nebelsick et al 2012). The beetle known to fog bask, Onymacris unguicularis, exhibits a decrease in roughness from the posterior of the elytra (where there are grooves) to the anterior of the elytra (where there are no grooves and the elytra is smooth). However, water runs from the posterior of this beetle's elytra to the anterior due to the inclined fog basking stance.
The 'narrow-leaf syndrome' was discussed by Martorell and Ezcurra (2007). They found that fog interception efficiency increased for narrower leaves. A parallel is drawn between this and the findings of Nørgaard and Dacke (2010) from the fact that the smallest of the four beetles harvested the most fog water and the later research by Nørgaard et al (2012), where it was suggested that a thinner similar boundary layer could explain the fact that a metal wire harvested a similar amount of fog water to that of the bushman grass. This could well be a point for future consideration with regard to a man-made fog harvester.
4.4. Hexagonally arranged microstructures
The SEM images taken of the three lizard species (Moloch horridus, Phrynosoma cornutum, Phrynocephalus arabicus), were observed by Comanns et al (2011) to be a honeycomb-like structure and they suggested this to lead to the hydrophilic nature of lizard skin. The beetle Physosterna cribripes also displays similar hexagonally arranged structures, although the arrangement on the beetle's elytra appears to be the opposite to that seen on the lizard's skin as it has a raised mid-section and the joining hexagonal array is depressed (which can be observed in Parker and Lawrence's paper). The SEM images of Physosterna cribripes taken by Guadarrama-Cetina et al (personal communication) study reveals honey-comb structure in the elytra valleys.
According to Comanns et al (2011), this hexagonal honeycomb structure increases the roughness of the surface and also causes more water to condense. With regard to the beetle elytra under investigation, a hexagonal-like structure existing in the valleys is the reason that water nucleates quicker in those areas (Guadarrama-Cetina et al (personal communication). Consideration should also be given to the fact that this increased ability for water to nucleate in the valleys could be due to the general elytra structure being one of bumps and troughs that could cause cooler air in the valleys and thus have a potential impact on the nucleation of water in these cooler areas.
Both these hexagonal-like structures observed on the lizards and beetle could have more to do with energy efficiency rather than adaption to water management properties; however, although it was thought appropriate to mention it here for the purpose of this review paper these structures will not be considered convergent in nature.
4.5. The scale of the various water management structures
Pulling all the information on the relevant observed convergences together with regard to scale information, table 4 below groups the convergent structures (i.e. cones- and groove-like structures; the hexagonal structures are included for interest) together for ease of comparison across the different species.
Table 4. Structure information.
| Structure | Species | Description | Scale | Source |
|---|---|---|---|---|
| Cones | Uloborus walckenaerius (This spider's wetted web) | Knot | Diameter = 21.0 ± 2.7 µm | Zheng et al (2010) |
| Physosterna cribripes (beetle) | Cone-like bumps | Diameter = 0.5–1.5 µm | Nørgaard and Dacke (2010) | |
| Hexagonal | Physosterna cribripes (beetle) | Hexagonal | Diameter = 10 µm | Parker and Lawrence (2001) |
| Distance between bumps (i.e. valley) | 0.5–1.5 mm | Nørgaard and Dacke (2010) | ||
| Moloch horridus (lizard) | Honeycomb | Diameter ≈ 10–20 µm | ||
| Depth ≈ 5 µm | ||||
| Phrynosoma cornutum (lizard) | Honeycomb | Diameter ≈ 15–25 µm | Comanns et al (2011) | |
| Depth ≈ 5 µm | ||||
| Phrynocephalus arabicus | Honeycomb | Diameter ≈ 20–30 µm | ||
| (lizard) | Depth ≈ 1 µm | |||
| Grooves | Stipagrostis sabulicola (grass) | Grooves | Diameter = 30–80 µm | Roth-Nebelsick et al (2012) |
| Ridge | Diameter = 100–150 µm | |||
| Opuntia microdasys (cactus) | Base grooves | Ave. width ≈ 6.8 µm | Ju et al (2012) | |
| Tip grooves | Ave. width ≈ 4.3 µm | |||
| Second level grooves along entire spine | Width ≈ 0.6 µm | |||
| Moloch horridus (lizard) | Grooved channel | 20 µm | Gans et al (1982) | |
| Uloborus walckenaerius (This spider's wetted web) | Joint | Diameter = 5.9 ± 1.2 µm | Zheng et al (2010) | |
| Onymacris laeviceps (beetle) | Elytra grooves | Width ≈ 0.1 mm | Nørgaard and Dacke (2010) | |
| Onymacris unguicularis | Elytra grooves | Width ≈ 0.5 mm | ||
| (beetle) | Separation = 0.1 mm |
It can be seen that all the grooves range from millimetres to micrometres, the hexagonal structures are all in the micrometre range with only limited scale information available from the literature reviewed with regard to the cone-like structures. Even though the cone-like structures given are in the micrometre scale, the scale of the cactus spine and the thorny cones on the lizard's scales will be in the millimetre range.
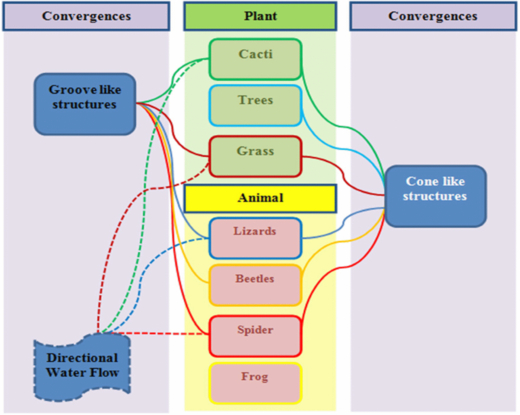
The identified convergences have been linked schematically in the illustration below (see figure 7). It can be seen that the groove and cone-like structures are not limited to plants or animals, they are abundant in both. Likewise, directional water flow was observed in both plants and animals. The hexagonal-like structures mentioned earlier were found only in animals and as such it is doubtful that they are convergent structures that aid the water collecting process.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 7. Schematic diagram linking nature's surface feature convergences and directional water flow.
Download figure:
Standard image High-resolution image{kind=link}
A study by Drabble and Drabble (1907) suggested that the absorption of atmospheric water is possible for plants that have a high osmotic leaf pressure; this could also be a point of consideration for future studies.
5. Implications and considerations
Humans have been utilizing dew, fog and rainwater for water collection for hundreds of years using nets, air well condensers, containers (e.g. water butts) and stones. It is only in recent years, however, that these fog nets and dew harvesters (i.e. passive, active and semi-passive) have made use of science, such as material science, to maximize and improve their efficiency. There has been particular focus on hydrophobic and hydrophilic properties with regard to fog harvesting nets, especially following the paper by Parker and Lawrence (2001). However, to date, nature's dew and fog harvesters have only minimally informed these fog-harvesting nets and dew devices and it is hoped that this review paper will start to influence the design of such devices, possibly with a focus on the convergences identified.
It appears that the animals and plants that have evolved in extreme conditions such as the hyper arid Namib Desert, with its extremely windy coastal fog, have adopted key water collecting features to survive. The surface irregularities that enable water to be pinned to the surface of the grass Stipagrostis sabulicola ensure harvested fog water droplets are not lost to the desert wind. It could be that observing the flow of wind-streamlines of moisture harvesting species will reveal some of the secrets of ensuring that fog water is not blown off the surface. When it comes to replicating these and future findings into something tangible for human use, one must always remember that the make-up of these species is due to several factors, such as protection from predators, and not just water harvesting.
Acknowledgments
This research paper was funded by Fujitsu and supported by HPC Wales and Swansea University. We would like to thank Jennifer Childs for casting her trained eye over the document for any obvious grammatical errors and SCIENTIA, the multidisciplinary research group at the Institute of Life Science for their support.
Footnotes
- 4
Laplace pressure (ΔP) is equal to 2γ/r, where γ is the interfacial tension and r is the interfaces curvature of radius. It thus follows, the smaller the conical radius (i.e. nearer a cone's tip), the larger ΔP. Thus the tapered radius from a cone's base to tip creates a Laplace pressure gradient across a droplet on a cones surface (Ju et al 2012).