





Abstract
The rapidly declining tropical forests of Asia support a diversity of felid species, many of which are rare and little known. We used camera traps in Dampa Tiger Reserve (TR), Mizoram, northeastern India, to estimate population density and describe activity patterns of 2 rare felids, the clouded leopard (Neofelis nebulosa) and marbled cat (Pardofelis marmorata). With a survey effort of 4,962 trap nights, we obtained 84 photo-captures of clouded leopards and 36 of marbled cats. We used spatially explicit capture-recapture methods to estimate population densities of both species. Using the Bayesian approach implemented in SPACECAP, we derived estimates of 5.14 (± 1.80 SD)/100 km2 for clouded leopards and 5.03 (± 2.07 SD)/100 km2 for marbled cats. Using camera-trap images, we compared diel activity patterns and activity overlaps for these 2 rare felids, together with 3 other sympatric carnivores, by estimating a coefficient of overlap between species. Among felids, clouded leopards and golden cats (Catopuma temminckii) displayed the highest overlap in activity, whereas marbled cats and leopard cats (Prionailurus bengalensis) showed the lowest, with marbled cats being primarily diurnal and leopard cats nocturnal. Our study provides the first density estimates from continental Southeast Asia for marbled cats and one of the highest recorded densities for clouded leopards. These results are of special significance since Dampa sustains an ecosystem that has in recent times undergone near extirpation of large predators.
Over 60% of global felid diversity is found in Asia, with 21 species occurring there (Macdonald et al. 2010). At least 11 of these species inhabit the tropical forests of South and Southeast Asia; approximately 40% of these tropical forests have already been converted to other land uses (Wright 2005), with the remainder being subjected to the highest rate of deforestation globally (Laurance 1999; Sodhi et al. 2004; Sodhi et al. 2010). An increasing human population (Sodhi et al. 2010), developmental pressures (Wilcove et al. 2013; Clements et al. 2014), hunting for subsistence, commercial (Corlett 2007; Velho et al. 2012), or traditional use (Chakravorty et al. 2011), and lack of effective and innovative protected area management (DeFries et al. 2005; Dickman et al. 2015) are among the issues threatening biodiversity and habitats in this region. Given these escalating pressures on felid species in Asia (Macdonald and Loveridge 2010; Dickman et al. 2015), both in terms of habitat loss and their value in commercial trade (D’Cruz and Macdonald 2015; Nijman and Shepherd 2015), a better understanding of their demography and behavior would be useful to inform conservation decisions.
The clouded leopard (Neofelis nebulosa) and marbled cat (Pardofelis marmorata) are felid species found in the tropical forests of South and Southeast Asia belonging to the Panthera (Johnson and O’Brien 1997) and Bay cat lineages, respectively (Johnson et al. 2006; O’Brien and Johnson 2007). Both clouded leopards and marbled cats are morphologically well suited for an arboreal lifestyle (Hunter and Barrett 2011) and are known to prey at least sometimes on arboreal species (Matsuda et al. 2008; Hunter and Barrett 2011). They are elusive, nocturnal, and found in low densities (Hunter and Barrett 2011; Nijman and Shepherd 2015). The clouded leopard is categorized as Vulnerable by the IUCN Red List (Grassman et al. 2015), while the marbled cat was recently relegated to Near Threatened from its previous status of Vulnerable (Ross et al. 2015). This change was primarily due to confirmed records of marbled cats from areas across its previously predicted distributional range with extensive use of camera traps in the region (Ross et al. 2015).
In 2007, clouded leopards were split into 2 species, N. nebulosa and N. diardi (Wilting et al. 2007). The former has a distributional range from Nepal in the west to Peninsular Malaysia in the southeast, whereas the latter is restricted to the islands of Sumatra and Borneo (Wilting et al. 2007). In the late 1980s, a series of surveys was conducted to determine the presence of clouded leopards in parts of Malaysian Borneo (Rabinowitz et al. 1987), Sumatra (Santiapillai and Ashby 1988), Taiwan (Rabinowitz 1988), Nepal (Dinerstein and Mehta 1989), and India (Choudhury 1993). While these data are important to map the distribution of the clouded leopard, information on their population densities and activity patterns is required to understand their ecological associations and to inform conservation policy. Given the charismatic appeal associated with clouded leopards (Macdonald et al. 2015), they constitute an important flagship species for tropical regions. The behavior of the marbled cat remains virtually unknown (Hunter and Barrett 2011), with the first population density estimate for the species being published from Sabah (Hearn et al. 2016), and nothing published on marbled cats across the continent (Ross et al. 2015).
Both clouded leopards and marbled cats are found in forested areas of northeastern India. They are generally considered rare and are encountered infrequently even in camera-trapping surveys (Datta et al. 2008). Their distribution stretches over remote areas with difficult terrain and political instability such as Manas Tiger Reserve (TR) in Assam (Borah et al. 2013), Intanki National Park in Nagaland, Namdapha TR in Arunachal Pradesh (Datta et al. 2008), and Dampa TR in Mizoram. While these factors cumulatively make clouded leopards and marbled cats difficult subjects of ecological studies, they also make them highly susceptible to extinction risks triggered by cultural and political reasons.
In this study, we estimated populations and assessed daily activity patterns of clouded leopards and marbled cats in Dampa TR in northeastern India. We also analyzed activity overlaps for these 2 species of felids and investigated their interspecific relationships with similar-sized sympatric carnivores. Previous studies indicate that an overlap of dietary and habitat preferences among similar-sized species results in competition, which can be avoided by temporal (Hayward and Slotow 2009; Lucherini et al. 2009) and spatial segregation (Fedriani et al. 1999). Hence, in this study, we predicted low temporal and spatial overlap between the larger clouded leopard and Asiatic golden cat (Catopuma temminckii) and the smaller, marbled cat and leopard cat (Prionailurus bengalensis). We include activity patterns of the dhole (Cuon alpinus) in our analysis since it is the most frequently encountered large predator in Dampa that shares dietary overlap with the clouded leopard, with a diet comprised of mammals such as red muntjac (Muntiacus muntjak) and wild pigs (Sus scrofa). We expect avoidance behavior between the 2 species due to both dietary overlap and the risk of interspecific killing of clouded leopards by dholes.
Materials and Methods
Study sites
We conducted this study between December 2014 and March 2015, in Dampa TR (23°20′–23°47′N, 92°15′ and 92°30′E) located in Mizoram state in northeastern India. Dampa TR is located in the Lushai Hills of the “Indo-Burma Biodiversity Hotspot” (Myers et al. 2000) and is an “Important Bird and Biodiversity Area” (BirdLife International 2015). The Chittagong Hill Tract region of Bangladesh is located to the west. The average elevation of Dampa TR ranges from 250 to 1,100 m asl (Raman et al. 1998). Annual average precipitation is 2,150 mm, received mainly from southwestern monsoons between May and October (Raman et al. 1998).
The dominant vegetation in valleys is comprised of evergreen species, whereas higher slopes have semi-evergreen or deciduous forest types (Raman et al. 1998; Birand and Pawar 2004). Until the early 1990s, large parts of the now core area of the reserve were under shifting cultivation regimes. These areas are now dominated by bamboo, comprised chiefly of Melocanna baccifera (Raman et al. 1998).
Dampa TR has a wide variety of mammals. Previous camera trapping conducted by the Forest Department in the Reserve to document biodiversity had revealed presence of both clouded leopards and marbled cats (L. Zathang, Mizoram Forest Service, pers. comm.). At least 30 species of mammals were photo-captured during our study. These include 4 species of felids (clouded leopard, marbled cat, Asiatic golden cat, leopard cat), 2 species of ursids (sun bear [Helarctos malayanus], Asiatic black bear [Ursus thibetanus]), and 2 species of canids (golden jackal [Canis aureus], dhole).
Field method for data collection
In April 2014, we conducted a preliminary reconnaissance survey in Dampa and Pakke TRs, located in northeastern India, in the states of Mizoram and Arunachal Pradesh, respectively. The survey was designed to identify an area of ~100 km2 in which to conduct a population estimation study of clouded leopards and members of their guild, using camera traps. During this period, we recorded locations of animal trails, felid scats and spoor, and locations with previous sightings or photo-captures of medium or small felids. This allowed us to identify our study site based on the logistic and administrative feasibility of camera trapping, assess security threats, and determine the ability to obtain adequate data on clouded leopards and other small felids.
Based on findings from the reconnaissance survey, we conducted camera trapping in an approximately 80-km2 area, located in the northeastern part of the core area of Dampa TR (Fig. 1). Apart from meeting other logistic requirements, in our opinion, this was the best-protected area of the reserve with relatively minimal human disturbance.
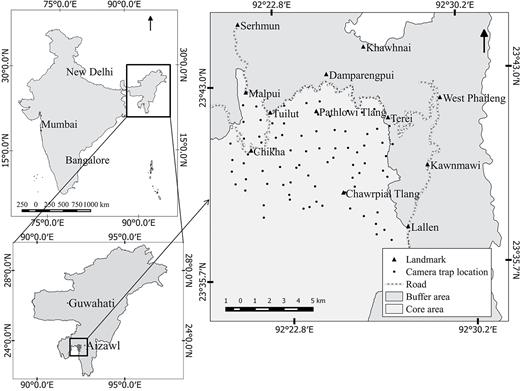
Map of the study area showing locations of camera traps (n = 79), road network around the protected area, and landmarks in the immediate vicinity of the area camera trapped. Landmarks included settlements and anti-poaching camps managed by the Forest Department within or around the study area. In the inset are maps of India and northeastern India, showing the approximate location of the study area.
At 79 locations, we stationed 148 paired units of Cuddeback Ambush IR camera traps (Model 1187; Cuddeback Digital, De Pere, Wisconsin). Each trap location had 2 camera traps placed on either sides of an animal trail facing each other, mounted 30 cm above ground. This arrangement allowed photographing both flanks of any felid that triggered the traps, and hence allowed greater reliability in identifying individuals using pelage patterns. Trap locations were identified based on presence of felid signs recorded during the reconnaissance survey, most of which were found on animal trails along stream beds and ridge lines. To optimize capture probability, we maintained a mean distance of 1.02 km (± 0.33 SD) between neighboring pairs of camera traps (Fig. 1). Previous studies indicated home ranges of 23 to 51 km2 for clouded leopards with exclusive core areas of 3 to 9 km2 (Austin 2002; Grassman et al. 2005a), whereas a marbled cat ranged over ~6 km2 based on few data points (Grassman et al. 2005a), hence we believe this intertrap distance was suitable for our target species. Camera traps were set to take single photos with the minimum delay time possible (15 s) between images to ensure we did not miss photographing 2 consecutive animals.
Traps were stationed at each location for a mean of 67 days (± 15.4 SD), and a maximum of 79 days. Some traps had to be removed sooner due to malfunction, elephant damage, theft, or security reasons (Supplementary Data SD1). Duration of our trapping period was less than that of other studies conducted on similar felids (Brodie and Giordano 2012; Wilting et al. 2012; Sollmann et al. 2014; Mohamad et al. 2015). Cameras were checked approximately every 2 weeks and batteries were changed once their levels reached 50%.
Analytical methods for population estimation
We used natural body and facial markings to identify unique individuals of both clouded leopards and marbled cats. We created individual encounter capture histories that used information in a binary format (detection or nondetection of the individual); along with other trap-specific information such as spatial coordinates of the trap, time, date, habitat type, and terrain. Each sampling occasion was comprised of a 24-h period.
To obtain density estimates, we used spatially explicit capture-recapture (SECR) methods that use spatiotemporal information associated with a capture. These methods eliminate the subjectivity associated with the calculation of an effective trap area to estimate density (Borchers and Efford 2008; Royle and Young 2008). All population density analyses were conducted in the R software environment (version 3.1.2—R Development Core Team 2015). We used the R packages “secr” (Efford 2015), which uses a maximum-likelihood-based approach (Efford et al. 2009), and “SPACECAP.” SPACECAP uses a Bayesian approach based on Markov Chain Monte Carlo method to fit capture data (Gopalaswamy et al. 2014) and is more robust with small sample sizes such as ours. We executed 100,000 iterations for both clouded leopards and marbled cats with a burn-in period of 10,000, thinning rate of 1, and an augmenting value of 220, which is > 10 times higher than the expected number of individuals (Gopalaswamy et al. 2012).
We created habitat suitability masks using the R package “secr,” with buffer sizes of 25 km for clouded leopards and 20 km for marbled cats, which are larger than those used for wide-ranging carnivores (Royle et al. 2009; Thapa et al. 2014). This mask is composed of suitable habitat for the study species, and along with the area covered by the trap array forms the “state-space” in SPACECAP. The state-space comprises a large number of equidistant points (in our study spaced at a distance of 580 m) that represent potential activity centers of the species being considered for the analysis and helps generate a pixel density map. We used large buffers to ensure that no individuals from beyond the study area were photo-captured (Chase Grey et al. 2013).
We modified the habitat masks for both species by clipping-out areas that we considered unsuitable to constitute home-range centers for either of our study species, resulting in suitable habitat of 2,534 km2 for clouded leopards and 1,735 km2 for marbled cats. Due to lack of information on habitat use by both target felids, we only considered areas occupied by human settlements as unsuitable habitat knowing that neither clouded leopards nor marbled cats would have the centers of their home ranges in such close proximity to human habitation.
Relative abundance index
In QGIS, we created a distance matrix between the protected area boundary and all trap locations. All traps were divided into 2 categories: 1) traps located within 1 km of the protected area boundary, called “Near Human-use Zone (NHUZ)”; and 2) traps located > 1 km from the park boundary, which we called “Away from Human-use Zone (AHUZ).” The distance for NHUZ was determined based on informal interviews with local community members on typical distances traversed by humans in search of forest produce around the forest boundary. We obtained a relative abundance index by calculating the number of photo-captures obtained for each species within a period of 100 trap nights (O’Brien et al. 2003). These were derived separately for both NHUZ and AHUZ. To obtain independent events, we followed the definition of independence used by O’Brien et al. (2003), where 2 photo-captures of the same species at the same camera-trap location were considered independent if: 1) they were consecutive photographs separated by a minimum interval of 30 min, or 2) they were consecutive or nonconsecutive photographs within a 30-min period, but where the individuals photo-captured could be identified as different individuals. This relative abundance index has been used in several studies in tropical forests (O’Brien et al. 2003; Datta et al. 2008; Farris et al. 2015) and acts as an index of species abundance allowing for easy comparison of data obtained across different study sites, especially in areas where the species capture rate is too low to conduct a population estimation study. In this study, while we cumulatively had adequate data to estimate populations for our study species in the study area, the lack of captures in the NHUZ did not allow us to determine population densities separately for the NHUZ and AHUZ, hence we chose to use the relative abundance index for comparisons between these 2 zones.
Analytical methods for daily activity pattern
To determine daily activity periods of clouded leopards and marbled cats and to better understand their temporal activity overlaps with other sympatric carnivores, we used the “overlap” package in R (Meredith and Ridout 2014). This method, while accounting for the circular nature of time, generates a coefficient of overlap for data from different species using a nonparametric approach for kernel density estimation (Ridout and Linkie 2009). The coefficient varies from 0 to 1, indicating complete temporal separation between 2 species at 0 and complete overlap at 1 (Ridout and Linkie 2009). To define a unique event, we used the same definition of independence used for the relative abundance index. On the basis of recommendations provided by Meredith and Ridout (2014), we used the estimator (Dhat4) for sample sizes more than 75 and the estimator (Dhat1) for smaller sample sizes, with 10,000 bootstraps for increased precision for each species analyzed and 95% confidence intervals (CIs) for the results generated.
Results
Our total survey effort was 4,962 trap nights from 74 pairs of camera traps; at 5 trap stations we obtained 0 trap nights due to loss of traps to theft or elephant damage (Supplementary Data SD1). We obtained photographs of 30 mammalian species during this period. Chiropterans and murids were exempted from the count because we could not identify them to genus or species levels.
Density estimation
We obtained 84 photo-captures of clouded leopards and 36 of marbled cats (Table 1). However, if the same individual was photo-captured multiple times on the same occasion by the same detector, it was treated as a single event, reducing clouded leopard captures to 81. Twenty pictures of clouded leopards and 14 of marbled cats were of poor quality and could not be used for individual identification. We identified 10 clouded leopards from 61 pictures and 10 marbled cats from 22 pictures (Table 1). Maximum recaptures for a single individual were 25 for a clouded leopard (Supplementary Data SD2a) and 7 for a marbled cat (Supplementary Data SD2b).
Summary of clouded leopard (Neofelis nebulosa) and marbled cat (Pardofelis marmorata) photo-captures obtained in northeastern India, December 2014 to March 2015, and used in the population estimation analysis.
| Clouded leopard | Marbled cat | |
|---|---|---|
| Total photo-captures | 84 | 36 |
| Total no. of unique individuals identified | 10 | 10 |
| Total number of photos used in the analysis | 61 | 22 |
| No. of pictures where an individual was photo-captured multiple times at the same detector on the same sampling occasion | 3 | 0 |
| No. of photos not used in the population analysis due to poor quality | 20 | 14 |
| No. of unique trap stations where species was captured | 23 | 8 |
| Clouded leopard | Marbled cat | |
|---|---|---|
| Total photo-captures | 84 | 36 |
| Total no. of unique individuals identified | 10 | 10 |
| Total number of photos used in the analysis | 61 | 22 |
| No. of pictures where an individual was photo-captured multiple times at the same detector on the same sampling occasion | 3 | 0 |
| No. of photos not used in the population analysis due to poor quality | 20 | 14 |
| No. of unique trap stations where species was captured | 23 | 8 |
Summary of clouded leopard (Neofelis nebulosa) and marbled cat (Pardofelis marmorata) photo-captures obtained in northeastern India, December 2014 to March 2015, and used in the population estimation analysis.
| Clouded leopard | Marbled cat | |
|---|---|---|
| Total photo-captures | 84 | 36 |
| Total no. of unique individuals identified | 10 | 10 |
| Total number of photos used in the analysis | 61 | 22 |
| No. of pictures where an individual was photo-captured multiple times at the same detector on the same sampling occasion | 3 | 0 |
| No. of photos not used in the population analysis due to poor quality | 20 | 14 |
| No. of unique trap stations where species was captured | 23 | 8 |
| Clouded leopard | Marbled cat | |
|---|---|---|
| Total photo-captures | 84 | 36 |
| Total no. of unique individuals identified | 10 | 10 |
| Total number of photos used in the analysis | 61 | 22 |
| No. of pictures where an individual was photo-captured multiple times at the same detector on the same sampling occasion | 3 | 0 |
| No. of photos not used in the population analysis due to poor quality | 20 | 14 |
| No. of unique trap stations where species was captured | 23 | 8 |
The closure test value for marbled cats (z = −0.21; P = 0.42) showed support for closure, and that for clouded leopards (z = −1.26; P = 0.104) weakly supported closure. Similar closure values are frequently obtained in studies of carnivores (Karanth et al. 2006; Soisalo and Cavalcanti 2006; Mondol et al. 2009). Our trapping period was shorter than that for most other recent studies on clouded leopards (Brodie and Giordano 2012; Wilting et al. 2012; Sollmann et al. 2014) increasing the likelihood that the assumption of closure is valid.
Using the Geweke diagnostic scores, we assessed for a convergence of simulations in our SPACECAP analysis. For clouded leopards, with a 25-km habitat mask, the Geweke score converged with all parameters being ≤ modulus 1.6 with z-score values of λ0 = 0.87; σ = 0.00; ψ = 0.11; Nsuper = 0.10; and Bayes P-value = 0.73. For marbled cats, the simulations converged with a 20-km habitat mask with z-score values of λ0 = 0.32; σ = −0.70; ψ = −0.36; Nsuper = −0.36; and Bayes P-value = 0.54. For marbled cats, even though the posterior mean density remained similar with a larger habitat mask of 25 km, we chose to use the 20-km habitat mask to ensure selection of the best-fit model for density generation. The posterior mean density estimate for clouded leopards was 5.14 (± 1.80 SD, 95% HPDI = 2.05–8.72)/100 km2 and that for marbled cats was 5.03 (± 2.07 SD, 95% HPDI = 1.67–9.50)/100 km2. These results were comparable to those obtained using the null model of the maximum-likelihood-based SECR method. Given the limited sample size of the data that we obtained, we could not use complex models that incorporate heterogeneity or covariates and hence restricted our analysis to the basic null model.
Through SPACECAP, we also obtained a pixel density gradient for the area, which provided a visualization of the spatial distribution and relative densities of the 2 species in the study area (Fig. 2). The pixel density gradients suggest high presence of both clouded leopards (Fig. 2A) and marbled cats (Fig. 2B) in the forested area between Serhmun and Tuilut. Another area with high densities of clouded leopards was south of the IR Police Camp at Chikha. The Chawrpial Tlang ridge line had a high density of pixels for marbled cats, although not so for clouded leopards, whereas a reverse trend was observed around Pathlowi Tlang, which showed high presence of clouded leopards but not marbled cats.
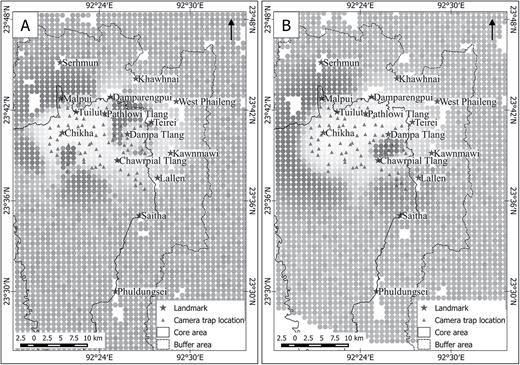
Maps of the study area and the surrounding landscape that constitutes the state-space in SPACECAP for A) clouded leopard (Neofelis nebulosa), and B) marbled cat (Pardofelis marmorata). Each of these surface density maps, generated by program SPACECAP, depicts the relative density of the 2 species for each pixel, sized 0.336 km2. Landmarks indicate settlements and anti-poaching camps managed by the Forest Department. The dots covering the map depict the relative pixel densities of each species, where the lightest shade of gray depicts the lowest relative pixel density and the darkest depicts the highest relative pixel density.
Relative abundance index
We obtained n = 391 independent photo-captures of the 5 primary predators in our study area (81 in NHUZ and 310 AHUZ). Similar spatial coverage of camera traps was maintained in both areas with a mean of 1.07 traps/km2 in the NHUZ and 1.02 traps/km2 in the AHUZ. The greatest difference observed in the index of relative abundance (number of photos/100 trap nights) between the 2 trapping zones was for clouded leopards (NHUZ = 0.60 and AHUZ = 2.33) and leopard cats (NHUZ = 0.71 and AHUZ = 2.05), with both species observed at greater abundance in the zones away from human use, whereas the least difference was for golden cats (NHUZ = 0.65 and AHUZ = 0.61). Marbled cats also showed similar relative abundance in both zones (NHUZ = 0.54 and AHUZ = 0.83). Dholes appeared to be using both zones (NHUZ = 4.09 and AHUZ = 1.91), but with greater relative abundance in the zone near human use.
Diel activity patterns and activity overlap
Temporal activity patterns (Supplementary Data SD4) and activity overlaps (Fig. 3) were obtained for 4 species of felids (n = 228) and 1 canid (n = 163). Clouded leopards showed their highest daily activity overlap (0.75, 95% CI = 0.60–0.86) with golden cats (n = 31; Fig. 3A) and their lowest overlap (0.63, 95% CI = 0.48–0.74) with marbled cats (Fig. 3C). However, while golden cats were relatively diurnal, with 67% of their photo-captures obtained between 0600 and 1800 h, only 40% of clouded leopard photo-captures were obtained during the same period (Supplementary Data SD3). Clouded leopards and leopard cats also showed relatively high activity overlap (0.73, 95% CI = 0.63–0.84; Fig. 3B). Two activity peaks (between 1800 and 0300 h and between 0600 and 1200 h) were observed for clouded leopards (Supplementary Data SD4A), suggestive of a bimodal activity pattern. The least-active period for clouded leopards between 1200 and 1800 h was synchronized with the highest activity period for marbled cats (Fig. 3C), with 47% of the photo-captures for marbled cats obtained within this period (Supplementary Data SD3).
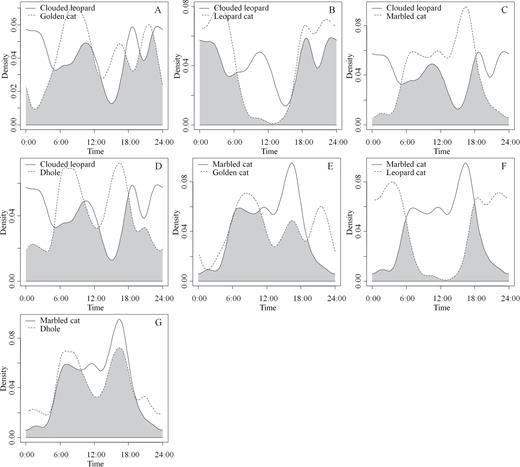
Overlap of activity patterns between sympatric predators in Dampa Tiger Reserve (TR), northeastern India, calculated using a kernel density function. The activity overlap between paired predators is depicted in gray. A) clouded leopard (Neofelis nebulosa) and Asiatic golden cat (Catopuma temminckii), B) clouded leopard and leopard cat (Prionailurus bengalensis), C) clouded leopard and marbled cat (Pardofelis marmorata), D) clouded leopard and dhole (Cuon alpinus), E) marbled cat and golden cat, F) marbled cat and leopard cat, and G) marbled cat and dhole.
Within felids, marbled cats shared their highest temporal overlap with golden cats (0.72, 95% CI = 0.58–0.88; Fig. 3E). The lowest daily activity overlap for any 2 species was recorded between leopard cats (n = 80) and marbled cats (Fig. 3F), with a kernel overlap density of 0.44 (95% CI = 0.25–0.51). Whereas 83% of marbled cat photo-captures were obtained between 0600 and 1800 h, 86% of leopard cat photo-captures were obtained between 1800 and 0600 h (Supplementary Data SD3), suggestive of strong nocturnal behavior for the latter species (Supplementary Data SD4D).
Dholes, although active at night, showed a strong bimodal pattern of activity, with activity peaks between 0600–0900 and 1500–1800 h (Supplementary Data SD4E). Dholes and marbled cats showed the highest daily activity overlap 0.81 (95% CI = 0.72–0.94) for any 2 species of top carnivores in the study area (Fig. 3G). A synchronized least-active period was noted between midnight and 0300 h for marbled cats, golden cats, and dholes (Supplementary Data SD3).
Discussion
Population estimation
We generated the first estimate of densities for marbled cats in continental Asia and an estimate using robust population estimation methods for clouded leopard populations in Dampa TR in northeastern India. Marbled cats found on continental Asia look different morphologically when compared with individuals from Borneo and Sabah and are rarely photo-captured by camera traps (Azlan and Sharma 2006; Datta et al. 2008), unlike the latter population, which appears to be relatively common (Hearn et al. 2016). Furthermore, given that forests devoid of tigers and other large predators are rapidly becoming common in mainland and Southeast Asia, this study holds high significance since it is typical of such places. Our density estimate of 5.14 (± 1.80 SD)/100 km2 for clouded leopards, using the Bayesian SECR approach, is comparable to the 4.73 (± 1.43 SE)/100 km2 estimate generated from Manas TR in India, using a maximum-likelihood-based SECR method (Borah et al. 2013). Our results indicate a high density of clouded leopards in Dampa compared to other parts of their range in Southeast Asia where the highest density obtained was 3.46 (± 1.00 SE)/100 km2 (Mohamad et al. 2015). Density estimates of the Sunda clouded leopard are generally even lower, with the highest density of 1.9 (95% CI = 0.7–5.4)/100 km2 reported from a primary forest in Sabah (Brodie and Giordano 2012).
We also obtained a relatively large number of photo-captures of marbled cats, which are believed to be primarily arboreal and very rare (Grassman et al. 2005a; Azlan and Sharma 2006; Lynam et al. 2013). The sigma parameter of 2,544 m (95% CI = 1,457–3,814) for marbled cats was similar to that for clouded leopards at 2,135 m (95% CI = 1,356–3,036), indicating that the 2 species traverse similar distances in Dampa TR.
The relatively high abundance of both felids in Dampa could be attributed to the local extirpation, or very low population density, of larger competing predators such as tigers (Panthera tigris) and leopards (Panthera pardus) as evinced by our camera-trapping study, in which no individual of either species was photo-captured. Although weak, a negative relationship has been recorded between clouded leopards and leopards (Ngopraserat et al. 2012), suggesting that the smaller clouded leopards may avoid the larger leopards. Another speculation is that smaller felids, in the absence of larger competitors, may become more terrestrial (Sunquist and Sunquist 2002), and hence more frequently camera trapped. Both these associations remain areas for further research.
Factors determining the spatial distribution of both clouded leopards and marbled cats also remain to be explored. In this study, a high presence of clouded leopards south of the IR Police Camp at Chikha (Fig. 2) could be an indicator of the camp acting as a deterrent to local hunters, subsequently resulting in higher prey presence (Jenks et al. 2012). However, the forested area between Serhmun and Tuilut, which shows high presence of both these species (Fig. 2), is relatively devoid of active surveillance by the Forest Department. On the eastern side of the study area, the 2 species show spatial segregation with high presence of clouded leopards around Pathlowi Tlang and high presence of marbled cats along the Chawrpial Tlang ridge line (Fig. 2). Past studies have shown that prey distribution (Szor et al. 2008; Ramesh et al. 2017) and habitat characteristics (Dickson et al. 2005; Szor et al. 2008; Ramesh et al. 2017) play important roles in distribution of carnivores, which we speculate also are important factors in the spatial distribution of these species.
Activity patterns
Carnivore abundances and distribution can be influenced by a multitude of factors, intraguild competition being an important one (Fedriani et al. 1999; Palomares and Caro 1999; Fedriani et al. 2000). Large carnivores, especially those living in social groups, can exert dominance over smaller-sized carnivores in terms of access to resources, kleptoparasitism, and interspecific killing (Palomares and Caro 1999; Linnell and Strand 2000; Hayward and Slotow 2009). This in turn could influence population abundances, distributions, and activity patters of sympatric carnivores. Our study revealed varying levels of activity overlap between all 4 felids found in Dampa (Fig. 3) with the clouded leopard and golden cat showing the highest temporal overlap, a behavior that has been previously reported for the 2 species from other parts of Southeast Asia (Lynam et al. 2013). Among the smaller-sized felids, expectedly, marbled cat and leopard cat showed the least temporal overlap (Fig. 3F) with the marbled cat being mostly diurnal (Supplementary Data SD4B) and the leopard cat being predominantly nocturnal (Supplementary Data SD4D), behaviors congruous to those reported from other parts of Southeast Asia (Azlan and Sharma 2006; Lynam et al. 2013). A high dietary overlap for leopard cats and marbled cats, comprised chiefly of small terrestrial and arboreal rodents (Austin 2002; Ross et al. 2015), could result in intraguild competition (Hayward and Slotow 2009) between the 2 species. We believe the temporal segregation between marbled cats and leopard cats observed in this study can be attributed to competition for similar-sized prey.
Although marbled cats and clouded leopards showed spatial overlap, as indicated by the pixel density gradients generated in SPACECAP, the 2 species exhibited low overlap in temporal activity, a behavior that has been recorded in other locations (Lynam et al. 2013). However, the separation in diel activity patterns of these 2 felids was not as distinct as reported by Lynam et al. (2013). Activity of clouded leopards was more nocturnal than diurnal, with a distinct period of low activity between 1200 and 1800 h (Supplementary Data SD4A). However, clouded leopards did show activity during the day, a behavior predicted for areas that lack larger predators (Van Schaik and Griffiths 1996).
We expected low temporal overlap between dholes and clouded leopards due to the risk of interspecific killing of clouded leopards by dholes. However, contrary to our expectations, the 2 species had relatively high (> 0.70) temporal overlap (Fig. 3D). We believe that clouded leopards may be able to avoid dholes by exhibiting arboreal behavior when they encounter the species (Austin 2002). Although clouded leopards and dholes both prey on medium-sized terrestrial mammals, competition for prey species in our study area also may be less than we predicted, with higher dependence of dholes on larger mammals such as sambar (Rusa unicolor). Further research on dietary composition and prey availability for both clouded leopards and dholes in the study area is needed to examine the influence of diet on activity.
As reported from other parts of their distributional range, dholes showed a distinctively bimodal temporal pattern with 2 activity peaks, in the morning and evening (Austin 2002; Grassman et al. 2005b). Although dholes were active primarily during the day, some activity was also recorded at night.
We believe, with lack of large predators and high prey availability, interspecific competition in Dampa may not be a critical factor influencing daily activity patterns of the existing carnivore guild. However, we acknowledge that these activity budgets have been generated only for terrestrial species using animal trails and streams beds. Our methodology would not detect arboreal behavior. Furthermore, these are grouped activity patterns for each species exhibited over large temporal and spatial gradients. Hence, we could not assess minor alterations in behavior based on perceived threats in a particular area or at a particular time.
Edge effect
We wanted to determine densities for clouded leopards and marbled cats separately for both NHUZ and AHUZ zones to gain a better understanding of edge effects on populations of these felids. The relative abundance index indicated that both clouded leopards and leopard cats had lower abundance in proximity to the park boundary, suggesting that the species may be avoiding areas along the park edge with relatively higher anthropogenic activities. The golden cat and the marbled cat used both the NHUZ and AHUZ zones similarly, which may be attributed to spatial displacement of the species induced by competition with clouded leopards. The relative abundance of dholes appeared higher in the NHUZ, which may be influenced by higher prey distribution in this zone, in proximity to human-use areas.
Limitations of the study
Our study provides the highest density estimates generated for both clouded leopards and marbled cats in mainland Asia. However, these estimates need to be considered with caution since they could be indicators of larger ecological processes underway in the region. The landscape around Dampa TR has undergone large-scale land-use changes in recent times. These include proliferation of oil palm plantations since 2007, encouraged by the state government under the “New Land-Use Policy” and horticulture promotion schemes (Mandal and Raman 2016), and expansion of the area under shifting cultivation with short fallow cycles (P. Singh, pers. obs.). Such changes in the surrounding landscape, although restricted only to the north and east of the study area, may have resulted in the recent concentration of these carnivores into the relatively undisturbed habitat in the core area of the reserve that forms our study area (Fig. 1). These carnivores may also respond differentially to the hard edges dominated by the expanding permanent monocultures of oil palm in the area and the soft edges marked by areas under shifting cultivation regimes, with resting periods when the forest can regenerate.
Although this study adds to our understanding of interspecific associations in our study area, these interactions may be better explained by a greater level of information on prey abundances and behavior (e.g., Karanth and Sunquist 2000). Overall, our study has revealed what may be one of the densest populations of clouded leopards and marbled cats in South Asia, while also providing insights into niche partitioning of their guild as reflected in activity patterns. We believe Dampa TR is an important area to protect rare, tropical biodiversity, including 4 species of poorly known felids.
Supplementary Data
Supplementary data are available at Journal of Mammalogy online.
Supplementary Data SD1.—Details of trap nights obtained at the 79 camera-trap stations within the study area in northeastern India.
Supplementary Data SD2a.—SPACECAP input file for clouded leopard (Neofelis nebulosa) data analysis. The file contains information of the detector at which a clouded leopard was photo-captured, identity of the individual photo-captured and the sampling occasion on which the event occurred.
Supplementary Data SD2b.—SPACECAP input file for marbled cat (Pardofelis marmorata) data analysis. The file contains information of the detector at which a marbled cat was photo-captured, identity of the individual photo-captured and the sampling occasion on which the event occurred.
Supplementary Data SD3.—Percentage of camera-trap photographs obtained of 5 sympatric predators during 3-h intervals in Dampa TR, northeastern India.
Supplementary Data SD4.—Diel activity patterns of major predators found in Dampa TR, northeastern India, shown using graphs that fit von Mises kernel density to time at which a species was photographed by a camera trap.
Acknowledgments
We express deep gratitude to Pu Liandawla, the Chief Wildlife Warden, Department of Environment and Forests, Government of Mizoram, for permission to conduct this study. We thank Pu Laltlanhlua Zathang and Pu Lalrinmawia, past and present Field Directors of Dampa, for administrative and logistic support. We are grateful to Ecosystems-India, and Nandita Hazarika for administering and co-ordinating this study and to the support of Dr. A. Zimmermann of WildCRU and Chester Zoo. We acknowledge the help provided by Pu Vanlalrema ACF and Pu Vanlalbera RO Terei and are immensely grateful to D. Barman, K. Lalthanpuia, Maunkima, Kaolauma, Thangliana, and others for assisting in data collection. We thank J. Ross and A. Hearn for their advice while analyzing data. We also thank the Robertson Foundation and anonymous donors to WildCRU that funded this study, which is part of WildCRU’s range-wide program on the clouded leopard guild.
Literature Cited
Author notes
Associate Editor was Bradley Swanson.

{kind=link}
{kind=link}
{kind=link}
{kind=link}