Evolution "for the Good of the Group"
By Edward O. Wilson, David Sloan Wilson
The process known as group selection was once accepted unthinkingly, then was widely discredited; it's time for a more discriminating assessment
The process known as group selection was once accepted unthinkingly, then was widely discredited; it's time for a more discriminating assessment

DOI: 10.1511/2008.74.380
Is evolution a team sport, or is the contest for survival played out strictly between individuals? There's no question that natural selection acts on individual organisms: Those with favorable traits are more likely to pass along their genes to the next generation. But perhaps similar processes could operate at other levels of the biological hierarchy.

In this way natural selection could perpetuate traits that are favorable not to an individual but to a social unit such as a flock or a colony, or to an entire species, or even to an ecosystem made up of many species. The underlying question is: Can biological traits evolve "for the good of the group"?
Many early biologists accepted the idea of group selection without thinking very critically about it. For example, herds of grazing animals might be described as evolving to conserve their food supply over the long term. Herds that exercised restraint would be more likely to survive than those that quickly exhausted a critical resource. Other biologists, looking more closely at such arguments, found a flaw in the reasoning. Prudently managing a shared resource benefits all members of a group, including any "cheaters" who consume more than their share. Genes associated with cheating would therefore spread through the group, and the propensity for cooperative resource management would be undermined. The situation is all too familiar in human experience; it is the phenomenon that Garrett Hardin famously named "the tragedy of the commons."
By the middle of the 1960s, ideas based on group selection were in deep disfavor, and the term itself was avoided in textbooks and the scholarly literature. When biologists observed behavior that appeared to benefit groups or species, they strove to explain it in terms of strictly individualistic selection. For example, animals might cooperate because they have genes in common ("kin selection") or because of the likelihood of reciprocal aid in the future. In this way, apparent altruism was interpreted as enlightened self-interest. It became almost mandatory for authors to assure their readers that group selection was not being invoked.
The time has come for a careful and forthright reassessment of group selection in evolutionary thinking. The most naive form of group selection—which axiomatically assumes that behaviors evolve for the good of the group—is clearly untenable. Nevertheless, traits with public benefits and private costs do evolve by natural selection. Just because cheaters have a within-group advantage doesn't mean that they prevail in the total population. Within-group selection is opposed by between-group selection, and the final outcome depends on the relative strength of these effects. Rather than categorically rejecting group selection and "for the good of the group" thinking, we need to evaluate the balance between levels of selection on a case-by-case basis.
To think clearly about group selection, it is important to compare the survival and reproduction of individuals in the right way. The problem with "for the good of the group" behaviors is that they are locally disadvantageous. A prudent member of the herd might gain from conserving resources, but cheaters within the same group gain even more. Natural selection is based on relative fitness. If solid citizens are less fit than cheaters within their own group, then something more is required to explain how they can evolve in the total population. That something is a positive fitness difference at a larger scale. Groups of solid citizens are more fit than groups of cheaters.
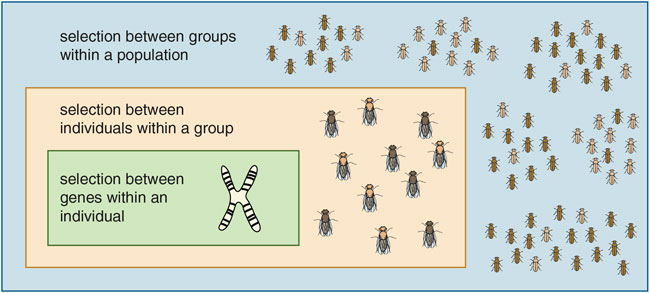
These interacting layers of competition and evolution are like Russian matryoshka dolls nested one within another. At each level in the hierarchy natural selection favors a different set of adaptations. Selection between individuals within groups favors cheating behaviors, even at the expense of the group as a whole. Selection between groups within the total population favors behaviors that increase the relative fitness of the whole group—although these behaviors, too, can have negative effects at a still-larger scale. We can extend the hierarchy downward to study selection between genes within a single organism, or upward to study selection between even higher-level entities. The general rule is: Adaptation at level X requires a corresponding process of selection at level X and tends to be undermined by selection at lower levels.
This way of thinking about evolution is called multilevel selection (MLS) theory. Although the term "multilevel selection" is newer than the term "group selection," the Russian-doll logic has been present from the beginning, going back to the works of Darwin.
Darwin would not have been motivated to think about group selection were it not for the existence of traits that are selectively disadvantageous within groups. In a famous passage from Descent of Man, he notes that morally upright people do not have an obvious advantage over less-upright people within their own "tribe," but that tribes of morally upright people would robustly outcompete other tribes. He concluded by saying "... and this would be natural selection." Darwin was clearly employing the Russian-doll logic of MLS theory in this passage. He did not comment on the irony that morality expressed within groups can become morally problematic in between-group interactions, but his hypothetical example perfectly illustrates the general rule stated above, which makes adaptations at one level part of the problem at higher levels.
Darwin's idea was elaborated by other evolutionists during the first half of the 20th century, notably the three founders of population genetics: Ronald Fisher, J. B. S. Haldane and Sewall Wright, who gave the idea a mathematical foundation. Their models differed in detail, but all embodied the Russian-doll logic of MLS theory. Unfortunately, many biologists were unaware of these models and thought naively about group selection, as we have already noted.
The events that led to the widespread rejection of group selection make an interesting story. In the late 1950s the University of Chicago was a hotbed of naive group selectionism. George C. Williams, a newly arrived postdoctoral associate, attended a seminar by Alfred E. Emerson, a termite biologist who likened all of nature to a termite colony. Williams was disgusted by the naivete of Emerson's claims and resolved to write a book clarifying the use and misuse of thinking about adaptation and natural selection. As Williams was writing, the Scottish biologist Vero C. Wynne-Edwards published a book titled Animal Dispersion in Relation to Social Behavior. Wynne-Edwards interpreted a vast array of animal social behaviors as adaptations to prevent the overexploitation of resources. He was aware that such prudence might be selectively disadvantageous within groups but assumed that it would evolve by group selection. Wynne-Edwards's overreaching claims were widely discussed and criticized. When Williams published his book Adaptation and Natural Selection in 1966, evolutionary biologists were in a mood to reach a consensus.
Williams forcefully asserted for a broad audience what a smaller group of experts already knew: Traits that are "for the good of the group" might evolve, but only by a process of group-level selection that is strong enough to overcome selection within groups. A key passage makes this point:
It is universally conceded by those who have seriously concerned themselves with this problem ... that such group-related adaptations must be attributed to the natural selection of alternative groups of individuals and that the natural selection of alternative alleles within populations will be opposed to this development. I am in entire agreement with the reasoning behind this conclusion. Only by a theory of between-group selection could we achieve a scientific explanation of group-related adaptations.
But Williams did not let the matter rest there. Having insisted that group selection is required to explain the evolution of group-level adaptations, he went on to claim that group selection is almost invariably weak, compared to selection within groups. In his own words: "Group-related adaptations do not, in fact, exist."
To summarize, Wynne-Edwards and Williams both accepted the Russian-doll logic of MLS theory and both made extreme claims that one level of selection routinely prevails over the other. Evolutionists found Williams more persuasive, leading to the widespread rejection of group selection. Generations of students were taught that group-level adaptations can evolve in principle, but do not evolve in practice, making "for the good of the group" thinking just plain wrong. Behaviors that appear to be for the good of the group must be explained in ways that are consistent with individual self-interest, such as kin selection or reciprocity.
In hindsight, it has become clear that both claims were too extreme. The balance between levels of selection can tilt in either direction. Between-group selection is sometimes a weak evolutionary force, as Williams supposed, but it can also be very strong, enabling groups to evolve into veritable superorganisms. There is no single formula; answers must be worked out on a case-by-case basis. Some examples of group-level adaptations will reinforce this point.
A good place to begin is with organisms that seem to blur the boundary between the individual and the group. For a spectacular example consider the elegant marine life forms called siphonophores, which include the Portuguese man-of-war. Many marine invertebrates live as colonies of individual organisms that are physically attached to one another. In some cases, such as corals, the members of a colony are undifferentiated and appear to function as autonomous units. A siphonophore, in contrast, is a colony made up of individuals with specialized forms and functions. Some members of the colony provide locomotion; others are charged with stinging and capturing prey; others do the work of digestion and assimilation. There is even a rudimentary nervous system. The siphonophores, in other words, have created a new kind of organism by turning simpler organisms into organs. Can their specializations be interpreted as "for the good of the colony" in the same way that organs can be interpreted as "for the good of the individual"?
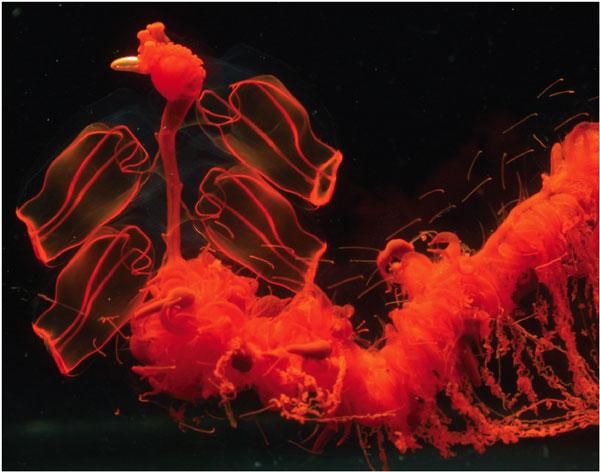
Most evolutionists would say yes, but it is instructive to review the reasoning that enabled an example such as this to be regarded as compatible with the rejection of group selection. Insofar as siphonophore colonies grow by asexual reproduction, their members are genetically identical. Of course the colony can be regarded as an adaptive unit, similar to a multicellular organism! What does this have to do with group selection?
To answer this question, we merely need to employ the Russian-doll logic of MLS theory. If all members of a colony are genetically identical, there can be no within-colony selection. Adaptations can evolve only by virtue of causing some colonies to survive and reproduce better than other colonies. The example represents a case of pure between-colony selection, not a denial of between-colony selection. Moreover, there is no warrant for assuming that members of a single colony—or the cells of a multicellular organism—are genetically identical. Mutations arise with every cell division, creating a potential for within-colony selection. Highly sophisticated adaptations evolved to suppress selection among genes and cell lineages within multicellular organisms and presumably also for siphonophore colonies. The bottom line is that division of labor and other design features of siphonophore colonies evolved by virtue of between-colony selection. If the example fails to impress, it is because it seems so obvious. Why, then, did evolutionary biologists accept the categorical statement that higher-level selection is invariably trumped by lower-level selection?
Another species on the borderline between individual and group is the cellular slime mold Dictyostelium discoideum. For most of its lifetime Dictyostelium consists of single-celled amoebae that forage in the soil, feeding on bacteria and other protists. When food grows scarce, thousands of amoebae come together to form a body called a slug, which migrates toward light over distances of up to 20 centimeters, then develops into a ball of reproductive spore cells held aloft by a nonreproductive stalk. Cells in the stalk eventually die, but the spores are dispersed (usually by sticking to a passing invertebrate) to start a new generation.
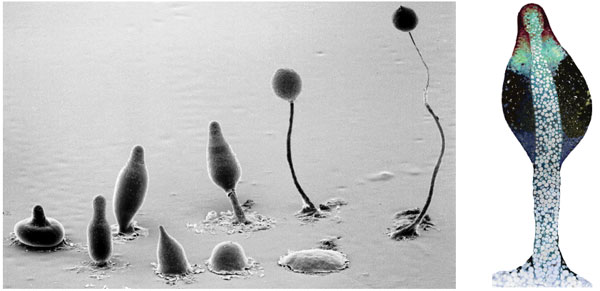
In the 1950s Dictyostelium became a model organism for the study of development, but the fact that the stalk cells appear to sacrifice their lives "for the good of the group" attracted little attention. Only later did Dictyostelium also become a model organism for the study of multilevel selection.
Before we take up the spectacular example of suicidal stalk formation, consider the subtler issues of slug formation, movement and orientation. Creating a slug requires the cells to secrete a polysaccharide matrix. Forward movement of the slug is accomplished by spiral motion of the cells within the matrix. Orientation toward light requires communication to coordinate movement. These are all examples of public goods that provide the same benefits for everyone but require individual effort. Even if the effort is nearly cost-free, as might be the case with orientation, the benefits are still shared (all individuals within a slug move to the same place), and so the fitness differences required to explain the adaptation reside at the group level (some slugs move to better places than others). There is no plausible way to explain these collective adaptations on the basis of within-group selection; they are group-level adaptations and can be accepted at face value as being for the good of the group.
Stalk formation appears to be the ultimate in self-sacrifice for the benefit of others. Individual cells have a powerful incentive to become reproductive spores rather than inert components of the stalk; and yet, if all the cells succeeded in becoming spores, there would be no stalk to promote dispersion. Selfish strains have been observed both in the laboratory and in the field. The problem is to show how between-group selection can be strong enough to counterbalance what appears to be an extreme selective advantage for selfishness within groups.
One possibility is that cell recognition enables amoebae to aggregate only with others that carry identical genes, so that each slug is genetically uniform. In this case natural selection would be purely at the group level, except for mutations, as with siphonophores. Another possibility is a lottery process analogous to the human social convention of drawing straws. In this case, some individuals make the ultimate sacrifice to become stalk cells, but they are a random sample, so there is no genetic selection within a slug. Current research indicates that neither of these explanations is entirely correct. Within-group selection does operate to a degree, favoring traits that are maladaptive at the group level. It would be just plain wrong to interpret these traits as being for the good of the group.
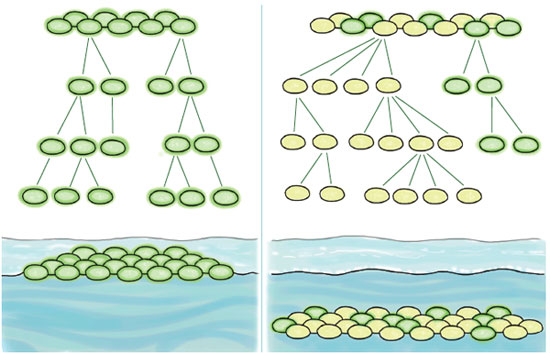
Some of the issues that arise in siphonophores and in Dictyostelium have also been explored in a laboratory setting in experiments with the bacterium Pseudomonas fluorescens. When this species is cultured in an unstirred broth, the cells soon consume most of the oxygen in the bulk of the medium, so only a thin layer near the surface remains habitable. A spontaneous mutation called wrinkly spreader causes cells to secrete a cellulosic polymer that forms a mat and helps them colonize the water surface. Production of the polymer is metabolically expensive, which means that nonproducing "cheaters" have the highest relative fitness within the mat; they get the benefit of the mat without contributing to its upkeep. However, if the proportion of cheaters grows too high, they are undone by their own success. The mat disintegrates, and the entire group sinks into the anoxic broth. Experiments by Paul B. Rainey and Katrina Rainey have shown that the wrinkly spreader trait is maintained in the population by group selection, even though it is disadvantageous within any one group.
The three examples described so far illustrate that between-group selection can be a significant evolutionary force and that group-level adaptations can be accepted at face value as "for the good of the group." Now let's use MLS theory to think about selection among genes within individual organisms.
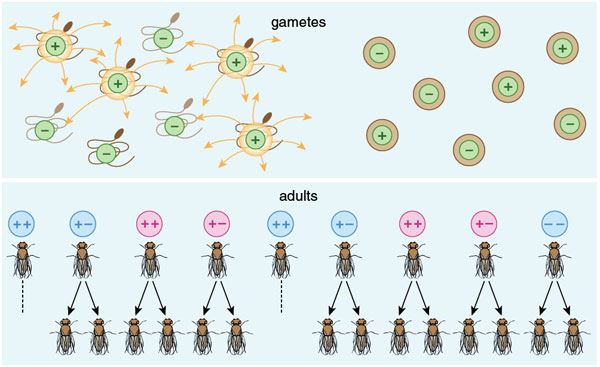
Meiosis is the reduction division that separates pairs of chromosomes in gametes. It is usually a fair process: The two genes at each locus on each chromosome have an equal chance of being represented in the next generation. The fairness of meiosis suppresses natural selection among genes within an individual and concentrates selection at the individual level (or above). That is why individuals are so functionally organized that they deserve the term "organism."
Some genes find ways to break the rules of meiosis, however, gaining a within-individual advantage through a process called meiotic drive. In the fruit fly Drosophila, a complex of genes known as segregation distorter, or SD, perpetuates itself even though it is detrimental to some of the flies that carry it. In males that are heterozygous for the SD genes, sperm cells bearing the genes secrete substances toxic to sperm lacking SD; as a result, almost all the surviving sperm are SD-positive, and the gene complex is overrepresented in the male fly's offspring. This is advantageous to the genes but not to the flies: Males that receive SD genes from both parents tend to be infertile.
This example shows how the Russian-doll logic of MLS theory can be applied to all levels of the biological hierarchy. Meiotic drive is "for the good of the gene" but it would be futile and erroneous to call it "for the good of the individual." It simply isn't. On the other hand, it is fully warranted to call an adaptation such as a turtle's shell "for the good of the individual" because it evolved by a process of selection among individuals. The same criteria that establish the legitimacy of "for the good of the group" thinking also establish the legitimacy of "for the good of the individual" thinking.
Our examples show how MLS theory can be used to evaluate the balance between levels of selection on a case-by-case basis, rather than declaring categorically that group selection always prevails (Wynne-Edwards) or never prevails (Williams). Why wasn't this judicious middle ground achieved long ago? The answer comes in part from a subtle shift in perspective that departs from the usual logic of MLS theory.
Consider the turtle's shell, a standard example of an individual-level adaptation. Genes that contribute to this adaptation are not more fit than alternative genes within any single turtle, but they are more fit than the alternative genes in the total population. That is just another way of saying that they evolve. Thus we can say that turtle shells evolve by gene-level selection, defined as the fitness of genes, all things considered. It is important to stress that nothing about the biological example has changed. The information for making the Russian-doll comparisons is still available; we are simply choosing to ignore the fact that the gene achieves its success by virtue of between-individual selection rather than within-individual selection.
The same gambit can be employed for individuals within groups. The wrinkly spreader strain is not more fit than the cheater strain within any mat, but it is more fit in the total population whenever mats evolve. In this fashion, all adaptations that evolve at level X, according to the Russian-doll comparisons, can be portrayed as lower-level adaptations. Ultimately, everything that evolves can be portrayed as adaptive at the gene level, regardless of where the fitness differences are located in the biological hierarchy.
This gene's eye view of evolution has always played a role in population genetics, where it is called "average effects" because it involves averaging the effects of alternative genes across all contexts to determine what evolves in the total population. Williams expounded on the concept in Adaptation and Natural Selection, where he aptly called it a "bookkeeping method." Most people, however, know average effects under the name "selfish genes," from the influential book of that title published by Richard Dawkins in 1976.
By itself, the gene's eye view interprets everything that evolves as adaptive at the gene level; however, even selfish-gene theorists need a way to distinguish a gene for meiotic drive from a gene for a turtle's shell. A concept called "vehicle of selection" is added to distinguish these cases. The individual is the vehicle of selection in the case of the turtle's shell because all of the genes in the turtle are "in the same boat" as far as their fitness is concerned. It should be obvious that the vehicle concept within selfish-gene theory duplicates the Russian-doll logic of MLS theory.
The point is that the Russian-doll logic is not arbitrary. It is essential for making sense of what we conventionally mean by "for the good of the individual" and, by extension, adaptations at all levels of the biological hierarchy. Numerous theories have been proposed to explain the evolution of apparent altruism and other solid-citizen behaviors without invoking group selection. When these theories are examined in detail, they are usually based on comparisons that depart from the Russian-doll logic of MLS theory. When reformulated in terms of the Russian-doll logic, they require group-level selection after all.
Nowhere has the pendulum of scientific thought swung more widely than in the study of eusocial insects (ants, wasps, bees and termites). Long before science existed as a cultural practice, honeybees were lauded as an example from nature of individuals acting for the good of the group. The great entomologist William Morton Wheeler was the first to describe eusocial insect colonies as "superorganisms" in 1911.
It might seem that if any biological phenomenon could survive the rejection of group selection, it would be the eusocial insects. Instead, eusocial insects became a particular focus of attempts to reformulate all evolutionary events as consequences of individual motives and acts.
The key to this program was kin selection, presented in the 1960s by W. D. Hamilton as an alternative to group selection. According to Hamilton's rule, an altruistic behavior evolves when br - c > 0, where c is the cost to the altruist, b is the benefit to the recipient, and r is the coefficient of genealogical relatedness, which ranges from 0 (for unrelated individuals) to 1 (for identical twins). When this inequality is satisfied, an altruistic act increases the absolute number of copies of the altruistic gene. For example, in some eusocial insects r is as high as 0.75, so that a cost of 1 is worth paying if the benefit to a colony-mate is at least 1.33.
Hamilton later realized that increasing the number of copies of an altruistic gene is not the same as increasing the fitness of altruists, relative to non-altruists in the same group. Working with another theoretical biologist, George Price, Hamilton reformulated his theory in the 1970s using the Russian-doll logic of MLS. To his surprise, he discovered that altruism is selectively disadvantageous within groups, even when the groups are composed of kin; it evolves in the total population only by virtue of between-group selection. His rule still predicted when altruism would evolve in the total population, but the theory was no longer an alternative to group selection; on the contrary, it required between-group selection. The coefficient of relatedness, which was originally interpreted as the probability of sharing genes identical by descent, could now be interpreted more generally as an index of genetic variation between groups. When r=0, the groups are randomly formed. When r=1, members of each group are genetically identical, and natural selection is concentrated entirely at the group level.
These developments should have led to the immediate revival of MLS theory in the 1970s, but intellectual momentum is not so easily reversed. From the present perspective it is clear that the vast majority of traits that enable social insect colonies to function as adaptive units can be accepted as products of between-colony selection. Some traits do evolve by within-colony selection, but these tend to disrupt colony-level functional organization.
One of us (E. O. Wilson) has described a social insect colony as "a factory inside a fortress." Both factories and fortresses suggest group-level functional design. The physical architecture of a termite mound, the caste system of an ant colony, the defensive response of a wasp nest and the foraging system of a honeybee colony are all communal activities that can be evaluated for their collective efficacy in the same way as the adaptations of individual organisms. For example, when a termite mound is damaged, workers rush to repair the breach, much as various kinds of cells repair a skin wound in an individual organism.
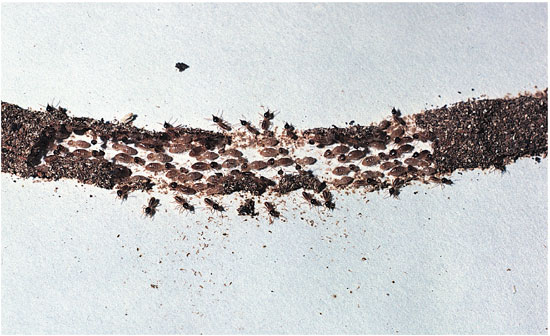
How did the termite behavior evolve? One possibility is that mound repair reflects within-colony selection: Individual termites with a genotype that predisposed them to make repairs survived and reproduced better than colony-mates with other genotypes. Far more plausible in this case is between-colony selection: Colonies in which the mound-repair genotype was common were more likely to survive and reproduce than other colonies.
Kin selection can be insightful in its own right, but it is profoundly misleading when interpreted as an alternative to between-colony selection. It also misleads us about the nature of variation among groups. Kin selection assumes that behavioral similarity is proportional to genetic similarity; the only way for a group to be behaviorally uniform, for example, is to be genetically uniform. However, colony-level adaptations in the eusocial insects usually rely on principles of self-organization, whereby relatively simple behavioral rules at the individual level interact to produce complex adaptive behaviors at the colony level. In self-organizing systems, small changes in the lower-level rules often produce large changes in the properties of the system as a whole. As a result, modest genetic variation among social insect colonies can produce substantial behavioral variation at the colony level, providing the raw material for colony-level selection. A correlation between genetic and phenotypic variation is required, but a high degree of genetic variation between groups (high values of r) is not. Even random genetic variation (r=0) can suffice.
Incipient social behavior is observed even in some solitary insects that do not share the peculiar kinship structure of the eusocial species. Bees in some species of the genera Certatina and Lasioglossum normally live alone, but when individuals are forced to live together, they spontaneously divide up tasks such as foraging, tunneling and nest guarding. Each bee is already programmed to perform these tasks on its own, switching among them as needed. But when a bee within a group encounters another bee that is already performing a task, it moves on to other tasks. Another pre-adaptation for social life is dominance behavior, which results in a prompt division of labor into reproductive and non-reproductive roles as soon as solitary bees are forced together. These adaptations in solitary insects that build and provision nests are "spring-loaded" to become adaptations for colonial life with minor modifications.
The concept of a group possessing a collective mind might seem like science fiction, but it follows directly from MLS theory and has been documented in impressive detail in the eusocial insects. Consider the parallel examples of a rhesus monkey making a decision and a honeybee colony making a decision. The monkey has been trained to watch a number of dots moving left or right on a screen and to turn its head in the direction that most of the dots are moving. Inside its brain, some neurons fire in response to right-moving dots and others fire in response to left-moving dots. A decision is made when one of these classes of neurons fires above a threshold rate. The honeybee colony has evolved to evaluate potential nest sites during the swarming phase of its life cycle. Higher-quality nest sites attract more scouts, and a decision is made when a threshold number of scouts is exceeded for one of the nest sites. The decision-making process is similar in the two cases, with individual bees playing the same role as monkey neurons.
Many other examples of distributed information processing—group minds—could be cited for the eusocial insects. Even more exciting, group minds are probably not restricted to the eusocial insects. As we have just shown, between-group selection does not necessarily require extreme genetic variation between groups. In the case of information processing, the collective benefits of making a wise decision can be great and the within-group costs can be low. Even individuals who are competing with one another can profit by making a wise collective decision about where to forage, providing a bigger pie to fight over. Evolutionary biologists are provisionally identifying group minds in species as diverse as African buffalo and American toad tadpoles. Theoretical models of consensus decision-making are often described in terms of self-interest because everyone in the group stands to gain from making a good decision. However, there are no fitness differences in a win-win situation. Employing the Russian-doll comparisons of MLS theory clearly reveals that collective decision-making evolves by between-group selection.
Group selection can easily be studied in the laboratory. One merely creates a population of groups and selects for a group-level trait, in the same way that individual-level traits have been artificially selected for centuries. These experiments almost invariably demonstrate a response to group-level selection. Often the magnitude of the response is greater than anticipated because of the way self-organizing systems amplify phenotypic variation.
William Muir of Purdue University compared two kinds of selection for egg productivity in hens. The hens were kept in cages, with several hens per cage. In the first experiment, the most productive hen within each cage was selected to breed the next generation (within-group selection). In the second experiment, all hens within the most productive cages were used to breed the next generation (between-group selection). In the first experiment, the most productive hen in each cage achieved her productivity largely by bullying the other hens. After six generations, a hyper-aggressive strain had been produced, with hens plucking each other's feathers in incessant attacks that were sometimes fatal. Egg productivity plummeted over the course of the experiment, even though the most productive hens had been chosen in every generation. In the second experiment, group-level selection resulted in a docile strain of hens, and egg productivity increased 160 percent in six generations.
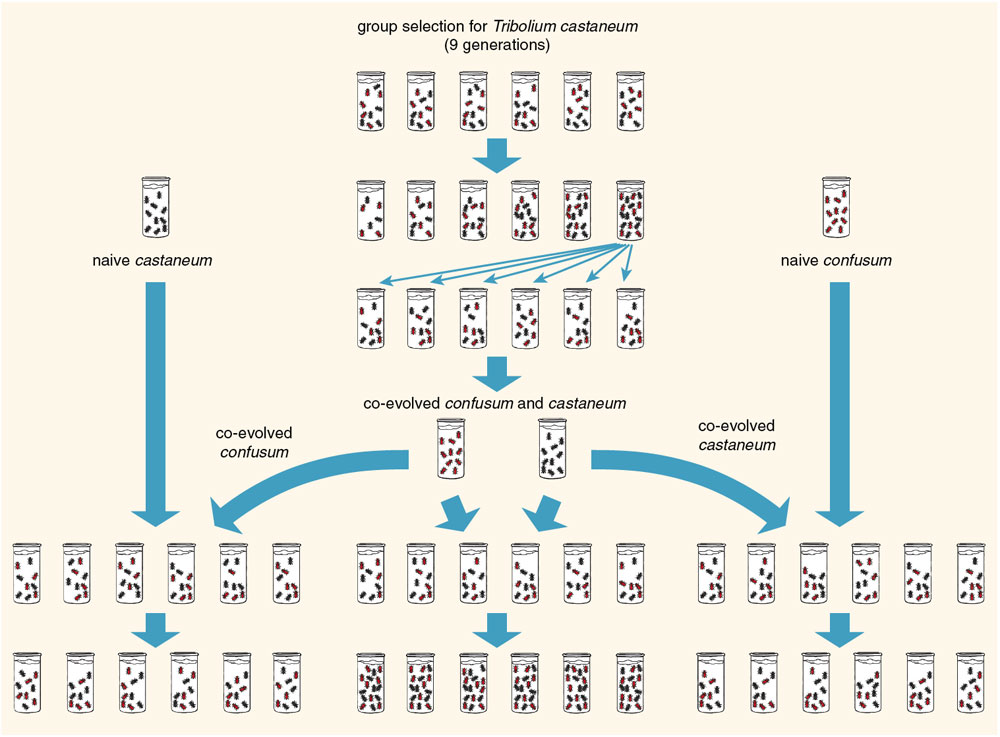
Laboratory experiments can also be used to explore selection at the level of multi-species communities. In the 1990s Charles Goodnight of the University of Vermont performed a series of ingenious experiments with two species of flour beetle, Tribolium castaneum and T. confusum. Goodnight placed equal numbers of beetles of the two species in vials and allowed them to interact and reproduce for several weeks. He then ranked the vials—each a two-species community—according to the population density of castaneum. Vials that scored highest were used as "parents" to create a new generation of vials. Even though the vials were selected based on the density of one of the species (the community-level phenotype), both species were transferred across generations as part of the same community, and each vial was started with equal numbers of each species. In other words, the unit of selection in the experiment was the whole community. Goodnight observed a response to community-level selection: The phenotypic trait (density of castaneum) increased over the course of the experiment, just as if he had been selecting an individual-level trait. In a second round of experiments, new communities were formed by combining the selected line of castaneum with confusum from the original stock population. In these reconstructed communities the evolved trait failed to appear. In this way Goodnight was able to show that community-level selection had produced genetic changes in both species, which interacted to increase the density of castaneum.
At a lower level of the biological hierarchy, an experiment like this one would be nothing out of the ordinary. If you breed Drosophila for a trait such as wing length, you can expect to see changes in genes on two or more chromosomes. In Goodnight's experiment the community plays the role of the Drosophila individual, and the two beetle species are like the interacting genes on separate chromosomes.
With William Swenson, one of us (David Sloan Wilson) has ratcheted the level of selection still higher in experiments with microbial ecosystems cultured in test tubes. Each test tube was inoculated with millions of microorganisms from a single well-mixed source (pond water), so that initial genetic variation between test tubes was negligible. Nevertheless, the test tubes quickly diverged from one another in measurable traits such as pH or ability to degrade a toxic substance. Both of these traits are properties of the physical environment influenced by living processes. Thus we were selecting a trait associated with an entire ecosystem, not with any individual or even any one species. Otherwise, the procedure was little different from selecting an individual-level trait such as wing length in Drosophila. Ecosystems that scored highest on the phenotypic trait were selected as "parents" to create a new generation of ecosystems. There was a response to selection, demonstrating that heritable phenotypic variation can exist at the level of whole ecosystems.
An unintended version of this experiment appears to have taken place in the selection of strains of yeasts and bacteria that produce kefir, a yogurt-like drink. In this case the ecosystem-level phenotypic trait is the taste of the kefir and its health benefits, causing some batches to be favored over others. Not only has this process selected a multispecies microbial community, but the community has evolved to aggregate into clusters held together by a polysaccharide matrix so that it can disperse across batches as a single unit.
These experiments raise the exciting prospect of creating "designer" groups, communities and ecosystems that perform useful functions such as degrading toxins, using artificial selection procedures that have been practiced on individual organisms for centuries. They also encourage us to look for examples of natural selection at the level of communities and ecosystems.
Individualistic theories of human behavior have been in vogue for the past half-century. Before then, it was common to think of human societies as being like single organisms. Indeed, the tradition of functionalism in the social sciences bears a strong resemblance to naive group selection in biology. Of course the uncritical application of "for the good of the group" thinking is no more justified in human affairs than elsewhere; between-group selection can prevail only when it is stronger than within-group selection. Having said this, however, we also assert that group selection has been a very powerful force in human genetic and cultural evolution.
Converging lines of evidence suggest that the key difference between human ancestors and other primate species was the suppression of fitness differences within groups, concentrating selection at the group level. Hunter-gatherer societies are fiercely egalitarian. Meat is scrupulously shared; aspiring alpha males are put in their place; and self-serving behaviors are censured. Unable to succeed at each other's expense, members of hunter-gatherer groups succeed primarily by teamwork.
Selection for teamwork probably began very early in human evolution. Human infants spontaneously point things out to others, and not merely to get what they want, which chimpanzees do not do at any age. Symbolic thought, language and the social transmission of information are fundamentally communal activities that rely on trustworthy social partners. Exploitation, cheating and free riding do exist in human groups, but what is most remarkable is the degree to which they are suppressed. They loom so large in our thoughts partly because we are primed to suppress them, like a well-adapted immune system.
Teamwork enabled our ancestors to spread throughout Africa and beyond, replacing all other hominid species along the way. While we remained a single biological species, we diversified culturally to occupy hundreds of ecological niches, harvesting everything from seeds to whales. The invention of agriculture added new layers to the biological hierarchy. We now live in groups of groups of groups.
When we confront the panorama of human genetic and cultural evolution, are we permitted to think about adaptations as being "for the good of the group"? As soon as we employ the Russian-doll logic of MLS theory, the answer becomes unambiguously yes. The idea that within-group selection invariably trumps between-group selection is as absurd for ourselves as it is for the eusocial insects.
Debates about adaptation and natural selection can become so contentious that they are called wars. In these wars, the most difficult ground to hold is the middle ground. In the 1970s Stephen Jay Gould and Richard Lewontin initiated a debate about the importance of adaptations compared to by-products of evolution ("spandrels") that became needlessly polarized. After the fighting was over, it was difficult to imagine what the fuss was about. Of course there are spandrels in addition to adaptations. Both must be identified on a case-by-case basis.
The controversy over levels of selection started earlier and is not yet quite over. In Adaptation and Natural Selection, Williams performed both a service and a disservice. The service was to forcefully assert that adaptation at any given level requires a process of selection at the same level and should not be invoked otherwise. The disservice was to adopt the extreme view that "group-level adaptations do not, in fact, exist."
We think that the time has come to declare a victory for the middle ground. Future evolutionists will look back and wonder what the fuss was about. Of course natural selection operates at multiple levels of the biological hierarchy. The Russian-doll logic of MLS theory must be used to evaluate the importance of each level on a case-by-case basis.
Click "American Scientist" to access home page

American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.