Integrating Ecosystem Resilience and Resistance Into Decision Support Tools for Multi-Scale Population Management of a Sagebrush Indicator Species
 Mark A. Ricca
Mark A. Ricca  Peter S. Coates*
Peter S. Coates*- U.S. Geological Survey, Western Ecological Research Center, Dixon, CA, United States
Imperiled sagebrush (Artemisia spp.) ecosystems of western North America are experiencing unprecedented conservation planning efforts. Advances in decision-support tools operationalize concepts of ecosystem resilience by quantitatively linking spatially explicit variation in soil and plant processes to outcomes of biotic and abiotic disturbances. However, failure to consider higher trophic-level fauna of conservation concern in these tools can hinder efforts to operationalize resilience owing to spatiotemporal lags between slower reorganization of plant and soil processes following disturbance, and faster behavioral and demographic responses of fauna to disturbance. Here, we provide multi-scale examples of decision-support tools for management and restoration actions that evaluate general resilience mapped to variation in soil moisture and temperature regimes through new lenses of habitat selection and population performance responses for an at-risk obligate species to sagebrush ecosystems, the greater sage-grouse (Centrocercus urophasianus). We then briefly describe general pathways going forward for more explicit integration of sage-grouse fitness with factors influencing variation in sagebrush resilience to disturbance and resistance to invasive species (e.g., annual grasses). The intended product of these efforts is a more targeted operational definition of resilience for managers by using quantifiable metrics that help limit chances of spatiotemporal mismatches among restoration responses owing to differences in engineering resilience between sagebrush ecosystem processes and sage-grouse population dynamics. Moreover, spatial resilience can be promoted though explicit consideration of sage-grouse and sagebrush predicted responses to active and passive management treatments across space and time. We describe tools that include multi-scale geospatial overlays and simulation analyses of post-disturbance land cover recovery aimed at prioritizing primary threats to sagebrush ecosystems in the Great Basin in the western portion of sage-grouse range (i.e., grass-fire cycles and conifer expansion), but underlying concepts have broader application to a range of ecosystems.
Introduction
Practitioners of restoration ecology continue to build upon the foundational concepts of ecological resilience (Holling, 1973), whereby pathways among ecosystem processes reorganize their structure following disturbances of various strength to either remain within an original state, shift among transient states, or fall into an alternative and possibly hysteretic state if thresholds for disruption are surpassed and return pathways are altered (Scheffer et al., 2001; Beisner et al., 2003; Suding et al., 2004; Standish et al., 2014). Arguably the largest impetus for this work is an increasing recognition of widespread changes to disturbance regimes, climate, and species pools occurring at local to global scales (Seastedt et al., 2008). These changes associate with a subsequent rise of novel ecosystems that are highly resilient against restoration efforts, owing to a deep and narrow basin of attraction in the alternative state, and are very difficult to manage (Hobbs et al., 2009). There is also now a greater appreciation of context dependency that seeks to recognize and identify the biotic and abiotic conditions that largely dictate the chances of restoration success (Eviner and Hawkes, 2008), and calls for more rigorous monitoring efforts of restoration outcomes with appropriate metrics over meaningful time periods (Suding, 2011).
Accordingly, managers, policy makers, and resource-user groups tasked with ecosystem stewardship during this challenging era of restoration (Suding, 2011) require tractable tools that bring resilience out of the conceptual realm and into effective implementation. The paradigm of “operationalizing resilience” has been proposed as means of achieving this daunting task and can be a powerful tool in the fight against permanent degradation and loss of vulnerable ecosystems world-wide (Suding, 2011; Angeler and Allen, 2016; Chambers et al., 2019a). Broad-scale efforts to make resilience concepts operational have occurred largely through socio-ecological frameworks such as the Millennium Ecosystem Assessment (Carpenter et al., 2009), the Resilience Alliance (2010), and Arctic stewardship (Chapin et al., 2015). However, their effectiveness is limited somewhat by a reliance on stakeholder-led iterative and qualitative processes rather than quantitative tools that predictively model outcomes of specific passive or active actions (Angeler and Allen, 2016). Thus, operationalizing resilience in restoration remains relatively nascent due to persistent challenges with quantification of factors influencing resilience under complex settings (Suding, 2011; Perring et al., 2015); and hence, the papers in this special issue. Another set of challenges toward operationalization are the multiple distinct, yet interconnected types of resilience that require clear definition to minimize confusion. Table 1 provides a brief description of the types of resilience invoked herein following definitions of Angeler and Allen (2016) and Chambers et al. (2019a).

Table 1. Brief definitions of types of resilience described for examples used in this paper following Angeler and Allen (2016) and Chambers et al. (2019a).
Operationalizing Resilience in Sagebrush Ecosystems
The iconic sagebrush (Artemisia spp.) biome of western North America spans 11 states and 2 provinces along varied hydrographic, floristic, and elevation gradients, harbors a diverse species assemblage of flora and fauna, and provides economic livelihoods for a diverse range of user groups including ranchers and outdoor enthusiasts (Suring et al., 2005; Davies et al., 2011). It is also a biome at risk, having contracted by over 50% post-European settlement (Schroeder et al., 2004) owing to multitude of factors including energy development, cropland conversion, improper livestock grazing, wildfire, and invasive species (U.S. Fish and Wildlife Service, 2015a). Concomitantly, over 350 plant and animal species occupying sagebrush ecosystems are of conservation concern (Suring et al., 2005; Davies et al., 2011), and none are perhaps more emblematic than the greater sage-grouse (Centrocercus urophasianus, hereafter sage-grouse). Sage-grouse are a well-documented obligate species to sagebrush ecosystems whose declining populations and threats to critical habitat have prompted multiple listing assessments under the Endangered Species Act (hereafter, ESA) since the end of the twentieth century (U.S. Fish and Wildlife Service, 2015a). Concern over the loss of sagebrush ecosystems and the broad ecological and socio-economic consequences of listing sage-grouse under the ESA spurred the development of science-based plans that lie at the nexus of one of the largest conservation efforts in United States history (Department of Interior, 2015; U.S. Fish and Wildlife Service, 2015a).
These efforts provided a strong catalyst for bringing well-studied factors influencing resilience to disturbance and resistance to invasive species (hereafter, R&R) into an actionable framework to help guide management decisions (Pyke et al., 2015; Chambers et al., 2017; Crist et al., 2019). To a degree, this was borne of a growing body of collective research that pointed to a frequent lack of sagebrush restoration success and resultant ineffective use of limited economic resources (Davies et al., 2011; Arkle et al., 2014). What rangeland managers needed was a tractable and spatially explicit tool that could predict baseline conditions associated with active and passive restoration success. Leveraging well-quantified linkages between soil moisture availability, primary productivity, and susceptibility to invasion that correlate strongly with elevation and are modified by aspect and vegetation-altering disturbances, Chambers et al. (2014) developed the foundational framework for predicting edaphic conditions associated with variation in R&R (in the sense of general resilience, Table 1) across sagebrush ecosystems. In brief, R&R is weakest at lower elevation sites characterized by Wyoming big sagebrush (A. tridentata wyomingensis) growing on warm and dry soil types, which are highly vulnerable to permanent transitions to novel and hysteretic ecosystem states driven by the loss of sagebrush, perennial grasses, and microbiotic soil crusts, and the subsequent domination of invasive annual grasses. In contrast, higher elevation sites characterized by mountain big sagebrush (A. t. vaseyana) and mountain shrubs growing on cool and moist soils where invasive annual grasses grow poorly have greater capacity for resilience. These concepts were made readily operational by classifying existing maps of soil climate that spanned the sagebrush biome into soil temperature and moisture regimes representing a gradient of underlying R&R properties (Maestas et al., 2016; Chambers et al., 2017). At its coarsest scale, soil regimes are aggregated into three categories that index R&R (low, moderate, and high), but use of temperature and moisture subclasses (Chambers et al., 2014, 2017, 2019a,b; Maestas et al., 2016) along with ecological site potential and collective responses to disturbance (Stringham et al., 2016) can facilitate applications with finer scale and grain (see section: Improving Estimates of Sagebrush Engineering and Spatial Resilience). This spatially explicit tool provided a foundation for triage of sagebrush management efforts across large spatial extents by identifying areas that would likely respond positively to active or passive restoration following disturbance vs. those that likely to respond poorly to restoration and hence prioritized for protection and management actions that enhance resilience (see section: Foundational Tools: Science Framework).
Threats to Resilience of Great Basin Sagebrush Ecosystems
Another important aspect of operationalizing resilience is understanding variation in biotic and abiotic stressors that provide energy for state changes across gradients of adaptive capacity that modify ecological resilience across large spatial extents (Gunderson, 2000; Scheffer et al., 2001; Folke et al., 2004). The Great Basin, comprising much of the western extent of the sagebrush biome, is larger than 80% of countries worldwide (Coates et al., 2016a), harbors > 45% sage-grouse leks rangewide (WAFWA, 2015), and includes an isolated Distinct Population Segment at the southwestern edge of the species' range (hereafter, Bi-State DPS) that has been evaluated separately for listing under the ESA (U S. Fish and Wildlife Service, 2015b). While sagebrush ecosystems and sage-grouse within the eastern portion of the range face threats arising directly from anthropogenic disturbances including cropland conversion and energy development (Doherty et al., 2016), managers of the iconic sagebrush ecosystems of the Great Basin face two primary biotic and abiotic stressors to R&R. The first is an accelerated cycle of wildfire driven by invasive annual grasses from Eurasia (hereafter, grass-fire cycle) and interactions with climatic conditions influencing loading and flammability of fuels, which is increasingly well-documented (Balch et al., 2013; Brooks et al., 2015; Coates et al., 2016a; Germino et al., 2016; Bradley et al., 2017; Pilliod et al., 2017a) and includes papers by Chambers et al. and Germino et al. in this special issue. Hence, we only briefly summarize the process here. This grass-fire cycle can be characterized by a non-analog positive feedback loop of fire, that kills most species of sagebrush, and is fueled by the invasion of winter annual grasses (mainly cheatgrass; Bromus tectorum) that can outcompete native perennial grasses by taking advantage of early fall and winter precipitation and high investment in seed production (Chambers et al., 2007). These grasses senesce by mid-late spring, much earlier than native perennials, and yield highly flammable fine fuels that spread fire to other stands of sagebrush that would otherwise not readily burn. The second is expansion of conifers, primarily single leaf pinyon (Pinus monophylla) and Utah juniper (Juniperus osteosperma), into otherwise treeless and sagebrush dominated communities, and driven largely by changes in land-use practices, past wildfire suppression, and apparent changes in climate post-European settlement (Miller et al., 2005; Romme et al., 2009). While more of a press than pulse disturbance (Bender et al., 1984) in comparison to the grass-fire cycle, conifer expansion over time concomitantly reduces dominance of sagebrush and perennial grasses (Miller et al., 2005), provides greater inputs of large wood to fuel more intense wildfires (Strand et al., 2013), alters faunal community composition (Davies et al., 2011; Donnelly et al., 2017) and hydrological function (Kormos et al., 2017; Stringham et al., 2018). The grass-fire cycle and conifer expansion collectively reduce general and ecological resilience through overall degradation of sagebrush ecosystem processes and excess fuel loading that increase wildfire probability, and spatial resilience (Table 1) through often hysteretic transitions to large grass or woodland dominated states that fragment otherwise continuous sagebrush.
Adding Sage-Grouse Metrics to Resilience Models for Sagebrush Ecosystems
Ecosystem restoration focuses largely on interactions between plants and soils given that they are primary determinants of productivity, yet failure to consider the response of fauna when planning and predicting restoration outcomes can be an obstacle against making resilience more operational (Perring et al., 2015). Lack of explicit attention to faunal response, particularly those at higher trophic levels, can stem from “field of dreams” concepts (Palmer et al., 1997; Sudduth et al., 2011; Perring et al., 2015) that assume both positive and rapid responses to restoration treatments providing habitat components necessary for life history demands. However, these treatments may not always yield resilient populations owing to spatial and temporal lags between reorganization of plant and soil feedbacks and corresponding demographic response of higher trophic taxa (Miller and Hobbs, 2007; Perring et al., 2015) (Figure 1). For example, lags can occur when population performance or generation times of higher trophic organisms progress too quickly relative to slower reorganization of plant and soil, or when changes to state factors such as climate and potential biota occur over large spatial extents or higher frequency such that spatial resilience is lowered by homogenization, even in ecosystems with relatively high resilience (Bestelmeyer et al., 2011). Consequently, if engineering resilience (i.e., rate of recovery to original condition, Table 1) is low, plant communities may eventually recover with or without active intervention over longer-time spans even after thresholds to disturbance have been surpassed, yet higher trophic organisms may show a more a hysteretic response (Bestelmeyer et al., 2011). Moreover, management actions intended to enhance ecosystem resilience to catastrophic disturbance and improve habitat quality for obligate species can create unintended ecological traps where animals select environmental cues that lower fitness (Battin, 2004).

Figure 1. Simple conceptual model illustrating asynchronies in engineering resilience between sage-grouse and sagebrush populations post-disturbance (e.g., wildfire). Even if sagebrush engineering resilience is high (as in high RandR habitats), sage-grouse population performance has lagged severely by the time sagebrush reaches pre-disturbance conditions. Reductions in survival and recruitment are illustrated by fewer hens with chicks; dashed lines represent time intervals; dashed horizontal lines represent closed populations. Spatial resilience is compromised when disturbance becomes widespread going from bottom to top.
It follows that while umbrella approaches focusing on single species have shortcomings (Andelman and Fagan, 2000), integration of metrics that account for measured responses of higher trophic taxa dependent on large and functional ecosystems should help facilitation of operational resilience (Suding, 2011; Perring et al., 2015). In our example, sage-grouse are well-recognized as an indicator species for the ecological integrity and conservation of sagebrush ecosystems at landscape scales, owing to the diverse array of community types used to meet life-history demands throughout their annual cycle (Rowland et al., 2006; Hanser and Knick, 2011; Runge et al., 2019). While not all ecosystem processes are covered completely under the umbrella of sage-grouse centric management approaches (Carlisle et al., 2018), evaluating resilience through additional lenses of sage-grouse habitat selection, population performance, and risks to persistence that are integrated with underlying sagebrush ecosystem R&R properties at multiple scales can help guide implementation and predict success of management actions (Chambers et al., 2017; Ricca et al., 2018). Herein, we: (1) summarize existing and new multi-scale tools, going from coarser to finer grain in terms of input data resolution and model complexity, as examples of integrating sage-grouse and sagebrush general and ecological resilience; and (2) describe general pathways forward for more explicitly integrating sage-grouse fitness and factors influencing variation in sagebrush R&R as metrics. In doing so, we aim to provide a more detailed operational definition of resilience for managers with quantifiable metrics that help guard against spatiotemporal mismatches owing to differences in engineering resilience between sagebrush ecosystem processes and sage-grouse population dynamics (Coates et al., 2016a), and how subsequent variation in feedbacks across space and time alter spatial resilience that contribute to sage-grouse population persistence across large spatial scales. We focus on tools aimed at addressing threats to sagebrush ecosystems in the Great Basin in the western portion of sage-grouse range, but the concepts presented have broader applications rangewide. Our examples are based largely on published model frameworks, so we direct interested readers to consult referenced papers herein for more information regarding specific methods and validations.
Foundational Tools: Science Framework
The Science Framework (Chambers et al., 2017; hereafter, Framework) serves as a solid baseline example for multi-scale integration of sagebrush ecosystem R&R concepts with ecological and management attributes associated with an indicator species represented by greater sage-grouse. Chambers et al. (2017, 2019b) provides a detailed summarization of the Framework and associated applications. Hence, a brief summary follows since it provides much of the conceptual basis of finer scale tools we describe next. A key element of the Framework at broad-to-mid scales is the spatially explicit intersection of data layers describing: (1) general resilience in sagebrush ecosystem R&R based on variation in soil temperature and moisture regimes described heretofore; and (2) a composite sage-grouse population index derived from lek-based models of sage-grouse breeding habitat probability and population abundance (Doherty et al., 2016). Importantly, incorporation of the sage-grouse population index represents an improvement over coarser-resolution available metrics such as percentages of sagebrush cover (Knick et al., 2013) or Priority Areas for Conservation (PACs, U.S. Fish and Wildlife Service, 2013) because it more directly accounts for habitat features selected by breeding sage-grouse in areas with abundant populations as determined by counts of sage-grouse at traditional breeding leks that are widely used to assess sage-grouse population trends (WAFWA, 2015). This property builds on the hierarchical approach of Coates et al. (2016b, 2019), which facilitates more precise prioritization of highly suitable habitats where sage-grouse are known to occur, while still accounting for unoccupied habitats of varying quality that may provide connectivity or other non-breeding life history needs. The binning of sagebrush R&R and sage-grouse population index layers into 3 respective classes each (i.e., high, moderate, and low) yields a 3 x 3 “sage-grouse habitat resilience and resistance matrix” that provides a highly tractable means for triaging management decisions relative to primary disturbance threats (e.g., conifer expansion, wildfire, invasive species) transcending broad to mid to local spatial scales across the species range (Chambers et al., 2016, 2017, 2019a,b). For example, reduction of conifer expansion in areas where shrub and herbaceous understories remain intact (i.e., Phase I or Phase II; Miller et al., 2005) can be aimed toward treatment of sites with underlying moderate to high R&R that are likely to support breeding sage-grouse. Wildfire prevention and suppression efforts are generally inversely related to R&R, and the strongest targeting occurs in high value areas characterized by low R&R that have high probabilities of breeding sage-grouse, where subsequent restoration efforts would have low chances of success. Moreover, the Framework facilitates ready inclusion of other spatially-explicit layers depicting relative risks of threats such as wildfire (Short et al., 2016), annual grass invasion (Boyte et al., 2019), and changing bioclimatic envelopes across different ecological gradients (e.g., Sage-Grouse Management Zones, Stiver et al., 2015). It can also readily adopt more complex models describing sagebrush ecosystem R&R, sage-grouse habitat selection and links to population performance, and disturbance threats at finer scale and grain; all of which aid effective targeting of management efforts to enhance operationalize resilience.
Examples of Increasing the Utility of Sage-Grouse Metrics to Operationalize Resilience Across Spatial Scales
Sage-Grouse Population Response to Sagebrush R&R-Based Recovery Models
Despite prolific seed production, other functional traits of sagebrush species dominating the Great Basin (e.g., mountain big, Wyoming big, black, and low) such as fire-induced mortality, slow-growth rates, lack of biotic and abiotic dispersal mechanisms, and high seed and seedling mortality (Pyke, 2011; Knutson et al., 2014; Schlaepfer et al., 2014; Shriver et al., 2019) hinder multiple types of resilience in the face of altered or novel disturbances, which includes the grass-fire cycle. Ecological and spatial resilience has been stressed by an increase in fire size, recurrence rates, and rotation intervals over at least the past 30 years (Brooks et al., 2015), which collectively provide more sustained energy to push heterogenous sagebrush communities into homogeneous cheatgrass-dominated states across large extents, particularly those with soil climates associated with low R&R that dominate (i.e., comprise over 50%) the Great Basin (Maestas et al., 2016). Both general and engineering resilience is influenced in part by the R&R gradient (Chambers et al., 2014), whereby differences in plant-available soil nutrients and moisture coupled with adaptive species traits drive variation in sagebrush growth rates and resistance to invasion following disturbances such as wildfire. Subsequently, spatial resilience is influenced by ecosystem responses to active and passive management that vary with R&R.
While building upon the basic premise of the Framework as it developed, Coates et al. (2016a) formulated a predictive and spatially explicit model that accounted for variation in these types of resilience following wildfire relative to underlying R&R conditions and ecological needs of sage-grouse, and related the output to demographic responses of sage-grouse across the entire Great Basin over 30 years. In brief, annual fire perimeters and severity indices (>1) obtained from the Monitoring Trends in Burn Severity database (Eidenshink et al., 2007) yielded spatially explicit data for both fire size and frequency of fire recurrence (i.e., reburning of previously burned area). These data were intersected with the soil-climate based R&R layer, and annual sagebrush recovery rates derived from previously published studies were assigned to R&R index classes. Relatively fast recoveries of 9 and 15 years were assigned to high and moderate R&R, respectively, and times were reset if fire recurred prior to recovery. In contrast, fire in low R&R were treated as permanent burn scars assumed to undergo a state-transition to cheatgrass. Importantly, engineering resilience post-fire was measured in terms of the amount to time necessary to provide a minimum of 20% sagebrush cover required for nesting sage-grouse. Sage-grouse population growth is sensitive to variation in nest survival (Taylor et al., 2012), and nest-survival is strongly tied to adequacy of concealment cover provided by sagebrush and other shrubs (Coates et al., 2017a). Sage-grouse also exhibit strong nest-site fidelity and do not readily vacate burned nesting habitat (Foster et al., 2019). Thus, the product of the model was an estimate of cumulative burn area (CBA) that accounted for chronic, rather than acute, wildfire effects on sage-grouse habitat needs during critical life-history periods relative to sagebrush recovery times that vary in relation to underlying R&R and fire recurrence rates. The amount of CBA has increased markedly over the last 30 years, with over 64,000 km2 affected as of 2016 (Figure 2). Moreover, even relatively rapid post-fire recovery of sagebrush to minimum thresholds of nesting cover in moderate and high R&R likely were not fast enough to overcome asynchronies with sage-grouse habitat needs, and further explained chronic effects of widespread wildfire on sage-grouse population growth (Figure 1). It also provided a powerful mechanism for explaining long-term declines of sage-grouse across the Great Basin, which simulated drought conditions by negating normally positive periods of population growth during infrequent years of above-average precipitation, and forecasting significant sage-grouse population declines through ~2040 even in habitats associated with high and moderate R&R if current rates of CBA remain unabated (Coates et al., 2016a). Moreover, the loss of spatial heterogeneity due to conversion of large swaths of burned sagebrush in low R&R areas, which dominate much of the Great Basin, to homogeneous stands of annual grass contributes to reduced spatial resilience and concomitant declines of sage-grouse with increasing CBA (Figure 2).
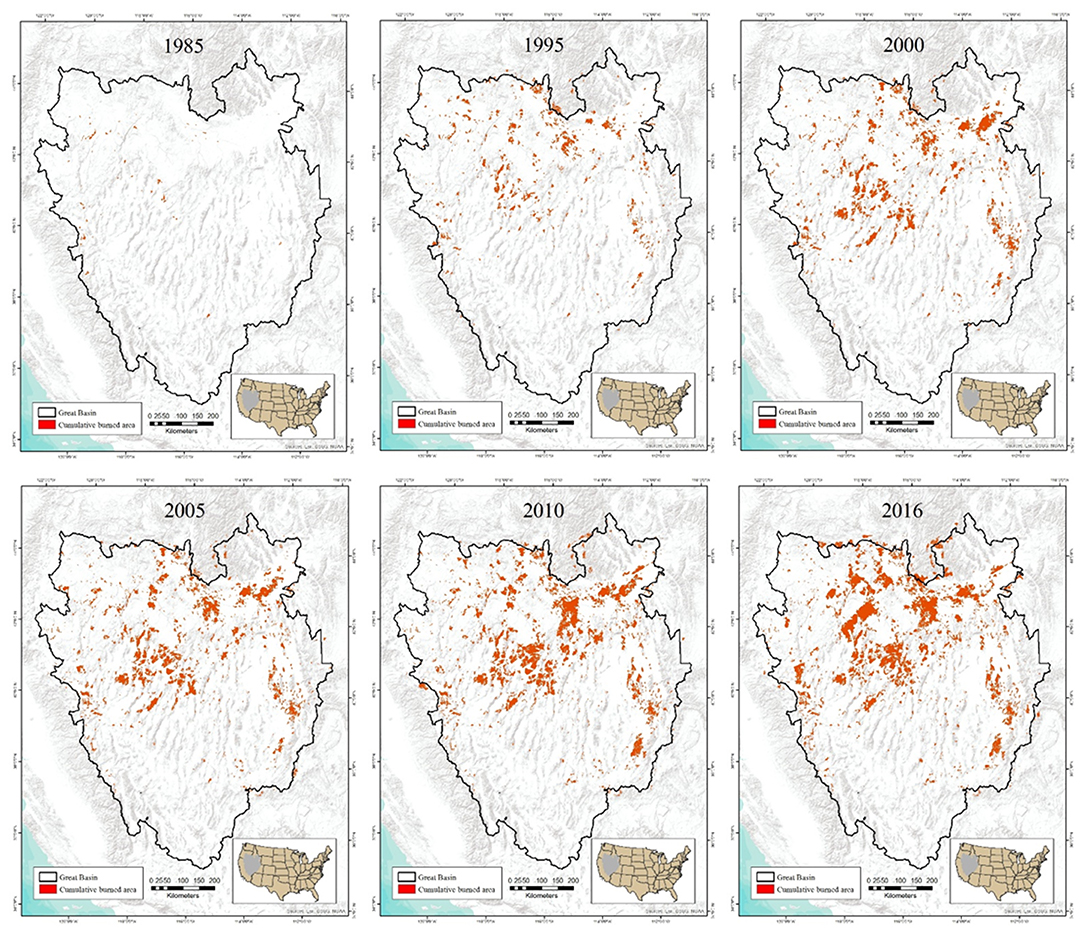
Figure 2. Time series depiction of cumulative burn area (CBA) at 5–6-year intervals from 1985 to 2016 modeled with RandR-based recovery rules from Coates et al. (2016a).
Refining Mid-Scale Spatial Intersections
A key component of the Framework is prioritization of management actions to prevent disturbance or identify best pathways for restoration following disturbance given underlying R&R and focal species (such as sage-grouse) needs at hierarchical broad, mid, and local scales. We provide three examples illustrating how finer resolution models depicting sage-grouse centric metrics can be integrated with R&R using the general geospatial overlay method of the Framework to address threats and prioritize management decisions at mid-scales (e.g., Great Basin, Bi-State DPS) stemming from wildfire and conifer expansion in a categorical fashion.
For our first example, we used a sage-grouse concentration area (hereafter; SGCA) geospatial layer modeled by Coates et al. (2016a) with the aim of identifying where wildfire management could be most beneficial to sage-grouse across the Great Basin. The SGCA was modeled as continuous surface using Doherty's et al. (2016) population index later integrated into the Framework, and then a threshold model was fit to identify where an increase in population index values no longer contributed to disproportionate population size relative to added habitat area. Binning the population index to this value (75%) identified areas that comprised <10% of the Great Basin but harbored nearly 90% of sage-grouse populations. Subsequent simulation modeling indicated that reducing the rate of cumulative burn area by 75% in SGCAs could halt declining rates of sage-grouse population growth (Coates et al., 2016a); hence, the SGCAs layer is ideal for use in geospatial exercises for operationalizing resilience to wildfire that follow. Spatially explicit recommendations for highest prioritization of wildfire management actions are then determined by intersecting burn probability (Short et al., 2016), CBA, SGCA, and R&R geospatial layers (Figure 3). Our overall prioritization scheme is similar to that described for the Framework (Chambers et al., 2016, 2017), but varies with respect to identification of finer scale SGCAs and past fire history that key in on sage-grouse centric metrics and estimates of sagebrush engineering resilience.

Figure 3. Example spatially explicit strategies for prioritizing wildfire management in the Great Basin based on geospatial intersections of fire probability (Short et al., 2016), wildfire impacted areas (Coates et al., 2016a), soil regime-based resilience and resistance indices (RandR, Maestas et al., 2016), and sage-grouse concentration areas (SGCA, Coates et al., 2016a). The top row represents example prioritization for wildfire prevention (A), suppression and initial attack (B), and restoration (C) given results of spatial intersections (lower row) for fire probability (left column) or cumulative (or recovered) burn area (middle and right column), and SGCA x RandR (middle row). Prioritizations are ranked in descending order.
Wildfire prevention areas are identified by intersecting the burn probability layer with the SGCA and R&R layers (Figure 3A). Priority for highest prevention can be placed in SGCAs likely to burn with low, followed by moderate, R&R. SGCAs with low R&R that are unlikely to burn can be afforded relatively less priority for prevention owing to limited fuel availability in these areas. Management actions such as conifer removal, targeted grazing, and strategic placement of fuel breaks for fuels reduction and staging areas for initial wildfire attack can be used to enhance prevention and resiliency to wildfire (Chambers et al., 2017, 2019b) in these priority areas. Once a fire ignites, spatial priorities for suppression and initial attack can be identified by intersecting the SGCA by R&R layer with CBA layer modified to illustrate recovered vs. non-recovered burned areas that as identified all past MTBS pixels with severity indices > 1 from the most recent annual CBA layer (Figure 3B). Highest priorities can be placed on SGCAs with low R&R that had not burned previously given low chances of active or passive restoration success and very low engineering resilience (Chambers et al., 2014, 2017). Subsequent high priorities can be placed in SGCAs with moderate and high R&R that had burned previously but recovered subsequently, followed closely by unburned SGCAs with moderate R&R. When prevention and suppression efforts fail, restoration resources can be expended in SGCA's with high and moderate R&R with recovered CBA, followed by the same areas that had not burned, given high chances of restoration success (Figure 3C). The rationale of prioritizing recovered CBA areas over previously unburned areas for suppression and restoration is predicated on efforts to reduce fire recurrence rates, and protection of SGCAs (and possible restoration investments) that likely now provide some minimal habitat requirements for sage-grouse after many years of sagebrush recovery and sage-grouse generations. Recovery processes can be further dissected by identifying burned high and moderate R&R pixels that have recovered vs. those in the process of recovery (Figure 4), and prioritizing SGCAs based on proportions of recovered CBA, recovering CBA, and unburned pixels. Prioritization rubrics can also be modified in instances, for example, where large unburned SGCAs are juxtaposed to smaller SGCAs with recovered CBA (Figure 4), or in burned SGCAs with low R&R where active restoration efforts show signs of success (e.g., Germino et al., 2018).
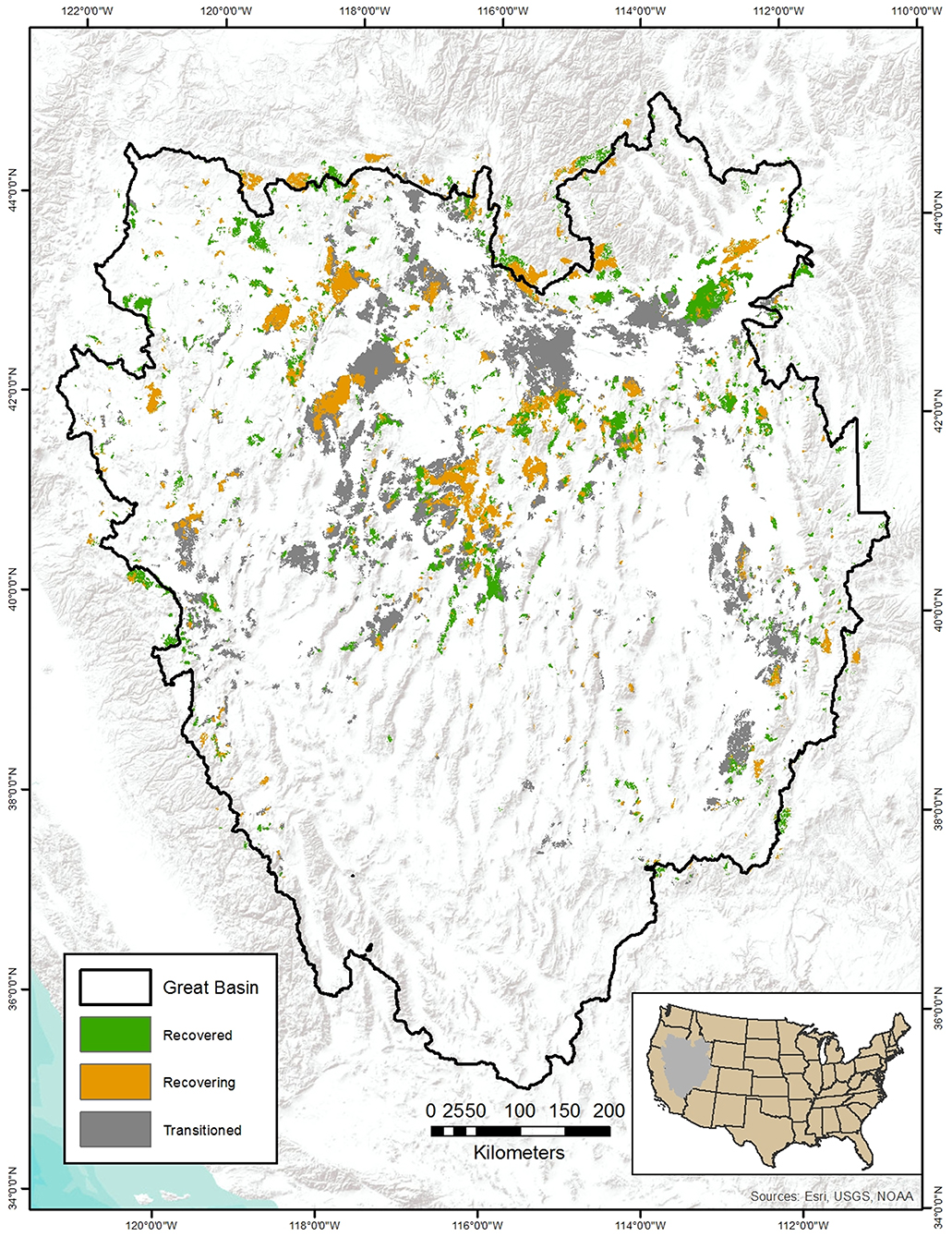
Figure 4. Alternative layer for depicting wildfire impacted areas in Figure 3 derived from geospatial intersections of high and moderate RandR areas with modeled complete or in-progress post-fire recovery, and areas of low RandR with modeled state-transition to annual grass using rules from Coates et al. (2016a).
Our second example builds upon the first, whereby we exchange the lek-based SGCA model at the ecoregion (i.e., Great Basin) extent with a telemetry- and lek-based model at the extent of Nevada and northeastern California to illustrate how inclusion of more localized models, where available, can provide finer resolution mapping of resilience-based management scenarios (Figure 5). For the latter case, habitat attributes disproportionately associated with leks relative to the random distribution of the same attributes provide more generalized models of habitat selection and hubs of population distribution because overall seasonal use patterns (particularly nesting) of non-migratory sage-grouse in the Great Basin are largely concentrated in diverse habitats within 5–8 km of leks (Coates et al., 2013; Manier et al., 2014). However, telemetry-based models can better account for specific resources selected differently among seasons by individual grouse to fulfill specific life-history needs such as nesting, brood rearing, overwintering, and movement corridors (Chambers et al., 2017). Compared to the SGCA-based overlay, a composite index derived by intersecting spatially explicit models of: (1) habitat selection informed by >44,000 locations from >1,700 telemetered sage-grouse that explicitly account for seasonal and regional climatic variation; and (2) a lek-based probabilistic index of abundance and space use (Coates et al., 2019) can provide finer scale depictions of predicted suitable habitat in areas likely occupied by sage-grouse (Figure 5). Subsequent intersections with R&R and recovered and non-recovered CBA layer using the same rules in Figure 3 for prioritization of wildfire prevention, suppression, and restoration can identify finer delineations for management actions such as fuel break and staging areas, tiered protection of continuous vs. fragmented habitat, and more targeted restoration efforts.
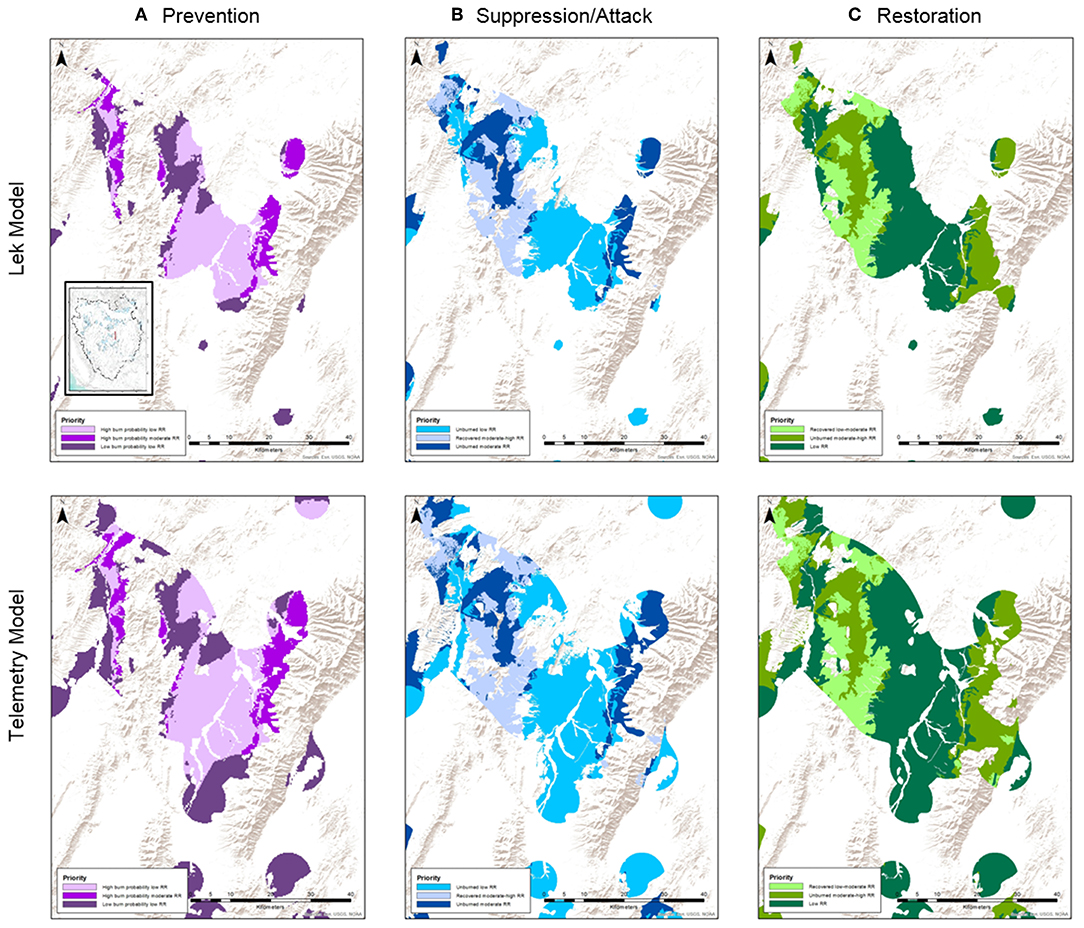
Figure 5. Illustration of finer resolution prioritization of wildfire management strategies using geospatial overlay of (Upper) a lek-only based model of sage-grouse habitat and abundance using SGCAs vs. (Bottom) a telemetry-and lek-based model of sage-grouse habitat and abundance.
Our third example illustrates how multi-scale areas for conifer removal can be prioritized by using categories derived from models that link sage-grouse habitat selection and concomitant impacts on survival to probabilities of conifer encounter and underlying R&R. Management efforts aimed at treatment of conifers expanding into otherwise treeless shrubland in the Great Basin through thinning or complete removal have accelerated greatly over the last decade (Severson et al., 2017a; Ernst-Brock et al., 2019). In addition to well-quantified effects of conifer expansion on sagebrush structure and function (Miller et al., 2005), these efforts have arisen from an increasing body of work quantifying how conifer expansion reduces sage-grouse population performance through decreased lek persistence (Baruch-Mordo et al., 2013), nest and brood survival (Sandford et al., 2017), annual survival (Coates et al., 2017b), and altered movement rates (Prochazka et al., 2017). While dense stands of continuous conifer woodland with depauperate understories (Phase III expansion, Miller et al., 2005) can be targeted for fuels reduction (Chambers et al., 2017), treatments for the benefit of sage-grouse populations are more commonly aimed at sparsely distributed trees in areas with dominant and intact shrub and herbaceous understories (Phase I), or to a lesser degree, in areas with higher conifer density becoming co-dominant with understories (Phase II). The Framework also suggests targeting of treatments in areas with high to moderate R&R to increase sage-grouse habitat selection and connectivity, as well as resilience to wildfire by reducing loads of heavy woody fuel (Chambers et al., 2017).
Restoration of habitat to fulfill life-history requirements for sage-grouse is readily accomplished in Phase I (and to a lesser extent, Phase II) owing to the need to remove relatively few trees and having an intact shrub and herbaceous component often requiring minimal reestablishment (but see Roundy et al., 2014). Rapid increases in cover of herbaceous vegetation can also ensue rapidly after treatment (Severson et al., 2017a), which correlate with increases in post-treatment population growth for sage-grouse (Severson et al., 2017b). Nevertheless, untreated Phase I encroached sagebrush can provide attractive resources to sage-grouse in terms of ample cover of shrubs and herbaceous vegetation. It follows that at the level of the individual, some sage-grouse demonstrate selection for areas of Phase I expansion, and likely do not perceive threats from low density trees contributing to increased mortality risk from raptors that perch and nest on trees (Coates et al., 2017b). These individual choices had significant fitness implications, whereby sage-grouse that demonstrated complete avoidance of Phase I had 20% higher survival probabilities compared to those individuals who demonstrated no avoidance. Reductions in survival were also most pronounced in areas of high R&R that corresponded to productive and mesic sage-grouse habitat, which also likely attracts raptors. Such areas could be significant ecological traps to individual sage-grouse owing to a decoupling of environmental cues that lead to maladaptive selection (Battin, 2004). Population-level impacts and concomitant reductions in spatial resilience of both sagebrush ecosystems and sage-grouse populations can occur if such traps are widespread, as is likely the case throughout much of the Great Basin. Moreover, deleterious impacts on sage-grouse lek persistence (Baruch-Mordo et al., 2013) and annual survival (Coates et al., 2017b) have been quantified at a threshold of 1.5–2.0% canopy cover, so treatments in both Phases I and II could have unintended consequences for sage-grouse if remaining canopy cover exceeds that threshold.
Spatially explicit delineations of conifer-associated ecological traps could help guide managers when prioritizing pinyon-juniper treatment areas within sage-grouse habitat. Accordingly, we use a geospatial overlay approach similar to that described heretofore, focused on the Bi-State DPS where conifers have been identified as a primary threat to sage-grouse and removal treatments are a key conservation tool (U S. Fish and Wildlife Service, 2015b) (Figure 6). We utilized a high-resolution (1-m2) map of conifer distribution and canopy cover derived from object-based image analyses of contemporary National Agricultural Imagery Program digital orthophoto quad tiles (Gustafson et al., 2017). Canopy cover classes (that index phases of encroachment) were estimated by calculating the proportion of mapped conifers within 900 m2 pixels, where cover classes 1, 2, and 3 represented >0–10, >10–20, and >20% conifer canopy cover, respectively. Cover class 1 was intersected with areas of high R&R to demarcate possible ecological traps (as in Coates et al., 2017b) (Figure 6A), We then intersected the same SGCA layer used in the wildfire example with the high R&R by cover class 1 layer to provide broad-scale targets for conifer removal in areas of abundant sage-grouse populations occupying selected habitat as measured from a lek-based model (Figure 6B). As with the wildfire example, telemetry-based models can be substituted to provide finer grain resolution. Here, we use a categorized resource selection function intersected with an abundance and space use index developed for the DPS (Ricca et al., 2018) and extracted to the 85% isopleth, which depicts finer grain across a large spatial extent (Figure 6C). We repeated the same approach using cover class 2 to demarcate areas where incomplete thinning efforts could lead to unintended ecological traps (Figures 6D–F). This approach can be expanded to larger spatial extents covering much of the Great Basin (Gustafson et al., 2017), and intersections with moderate R&R classes performed to further triage removal priorities outside of possible ecological taps that occur in selected and occupied habitat, but have higher chances on annual grass invasion following treatment. Risks of treatment vs. improvement in sage-grouse habitat should be weighed particularly in dry sites with low perennial herbaceous cover following mechanical treatment (Roundy et al., 2014; Bybee et al., 2016).
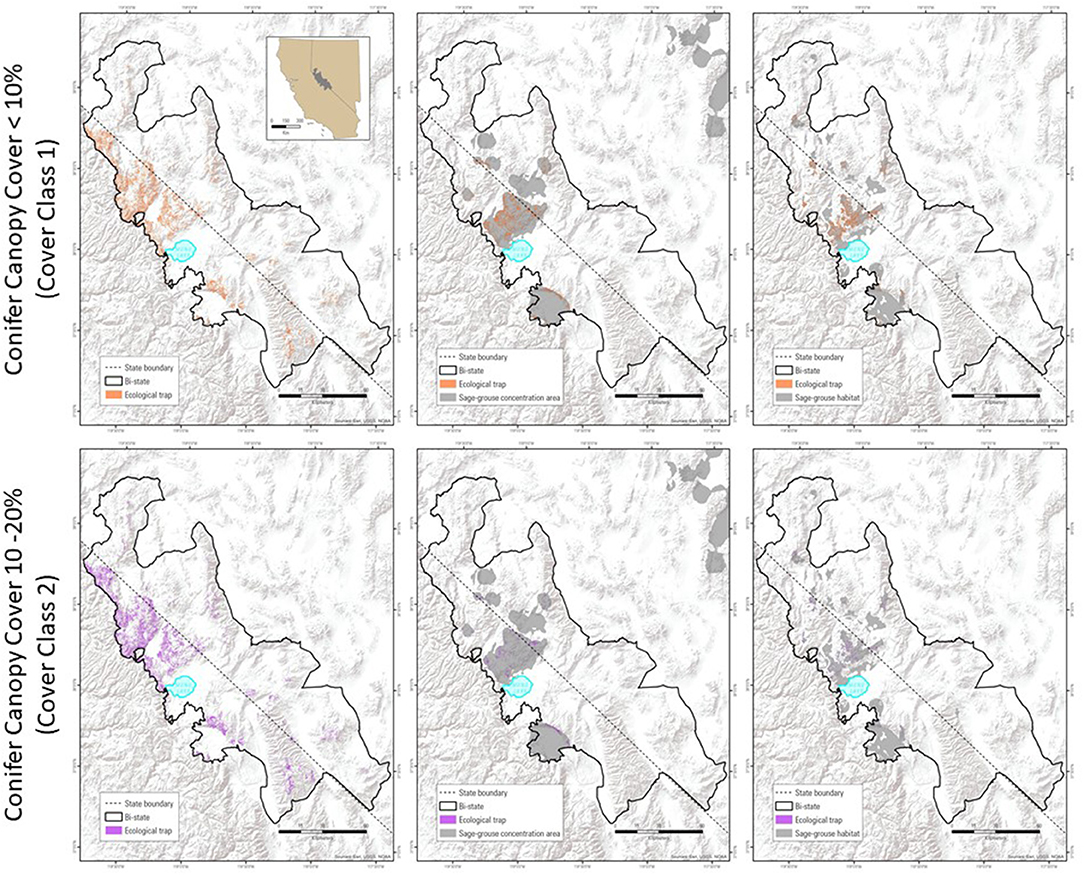
Figure 6. Spatial overlay approach depicting the distribution of possible ecological traps to sage-grouse based on the intersection of cover-class 1 conifers with high RandR (upper row, A), and further delineated for management priority based on intersections with a lek-only based model of sage-grouse habitat and abundance using SGCAs (B), and a telemetry-and lek-based model of sage-grouse habitat and abundance (C). The lower row (D–F) represents ecological traps that could be created by thinning of cover-class 2 to cover-class 1 in high RandR areas.
Scaling-Down Mid-Scale Models to Better Inform Local Site Selection Processes
The mid-scale spatially explicit models of sage-grouse habitat selection and abundance distribution intersected with predicted responses to disturbance (e.g., conifer expansion and wildfire) given underlying R&R properties help better identify areas for management across larger landscapes. Site-level implementation of management is also a key component of the Framework (Chambers et al., 2017; Crist et al., 2019), and direct application of mid-scale models may be too coarse in some cases to inform the most effective targeting of treatments given within site heterogeneity. In our examples, substantial variation in habitat selection within areas identified at the mid-scale can occur given inter-site differences in the availability of resources required by sage-grouse (Coates et al., 2019), and how that availability changes subsequent to disturbance and management carried out at a finer grain within the local scale. Here, we describe recently developed decision-support tools for conifer treatment and fire restoration that downscale mid-scale models, or leverage existing and extensive site-specific models, and apply simulated changes to land cover or habitat characteristics and concomitant quantified improvement in habitat quality to sage-grouse across candidate treatment sites while implicitly or explicitly considering underlying R&R. Such tools are also especially helpful when disturbance is widespread across mid-scale identified areas, but limited resources are available for uniform implementation of restoration treatments that are intensive and costly. In the process, spatial resilience can be enhanced by avoiding implementation of likely ineffective active management action within mid-scale identified areas.
Two recent studies (Reinhardt et al., 2017; Ricca et al., 2018) applied spatially explicit mid-scale models of sage-grouse resource selection across different life-stages to proposed or existing conifer removal treatment units identified by resource management agencies (e.g., Bureau of Land Management and Forest Service districts). Another recent study applied similar approaches to inform conifer removal for Gunnison's sage-grouse (Centrocercus minimus) but did not consider general R&R explicitly or implicitly (Doherty et al., 2018), so we excluded it from our review. In the Ricca et al. (2018) study, the goal was to rank candidate treatment units based on improvement of annual habitat selection following removal of Phase I conifer in the Bi-State DPS. In brief, a baseline resource selection function (RSF) describing features selected annually by sage-grouse of both sexes, including reproductive and non-reproductive females, status was calculated by contrasting existing land cover, topographic, and hydrologic attributes at multiple scales (e.g., moving windows) at used radio-telemetry locations compared to those at random locations. Conifer treatment and restoration of underlying shrub and herbaceous pixels were then simulated in a geographic information system (GIS) by: (1) removing conifer pixels, as measured from high-resolution mapping (Gustafson et al., 2017), comprising Phase I expansion (as indexed by cover class (1) within candidate treatment units, (2) returning understory pixels to their land cover type (e.g., big sagebrush, non-sagebrush shrub) without conifer overstory, and (3) re-running moving window analyses on land cover types and applying baseline RSF coefficients to the post-treatment landscape. Differences between pre- and post-treatments relativized RSF surfaces reflect per area increases in habitat selection, and multiplication by the lek-centric abundance and space-use index (AUI) account for spatial resilience by assigning higher rank to treatments in closer proximity to existing and sage-grouse populations (Figure 7). Final intersections with R&R allow inspection of highly ranked treatments compared to relative risks of annual grass invasion following disturbance from removal. Notably, this approach indicated that the majority of ecological benefits can be comprised within just a few treatment units, treatments of identical cost can have substantially different benefits to sage-grouse, and focusing solely on treatments in high and moderate R&R can disqualify the highly ranked treatments that occur in low R&R, yet they can be treated with relative low risk using non-mechanical methods (Bybee et al., 2016).
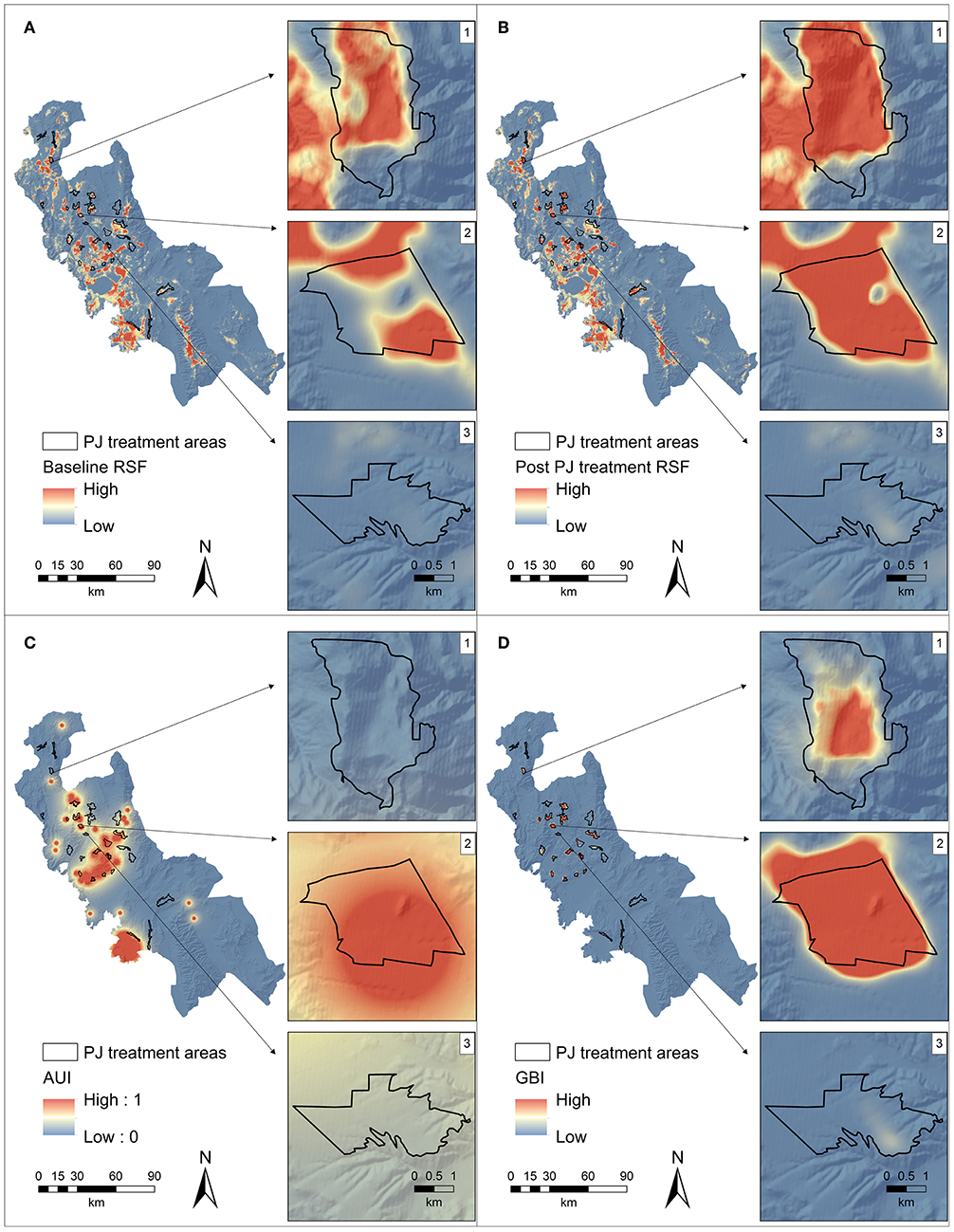
Figure 7. Example spatially explicit illustration of effects of simulated conifer removal and subsequent change in ecological benefit to sage-grouse to prioritize treatments. Here, resource selection function (RSF) values between baseline (A) and post cover-class 1 pinyon-juniper removal (B) surfaces are subtracted, and then multiplied by an intersecting abundance and space use index (AUI) (C) to calculate a sage-grouse benefit index (GBI) (D). Side panels illustrate how high GBI rankings can be driven by high RSF change and low AUI (1), high RSF change and high AUI (2), and low RSF change and moderate AUI (3). Used with permission from Ricca et al. (2018).
In the Reinhardt et al. (2017) study, the goal was to prioritize conifer removal efforts at the mid-scale (southeastern Oregon) through a process that optimized improvements in breeding habitat, movements between breeding and brood-rearing habitats, and inter-PAC movements. That process used an ensemble of models describing lek distribution, breeding sage-grouse habitat selection (Doherty et al., 2016), conifer cover (Falkowski et al., 2017), mesic habitats for brood-rearing (Donnelly et al., 2016), landscape resistance (Knick et al., 2013), and R&R (Maestas et al., 2016). Costs were factored as combinations of conifer cover and R&R class (whereby high cover and low R&R have the highest risk). Prioritized sites were characterized by low canopy cover, high R&R, and abundant sage-grouse populations. Moreover, spatial resilience was enhanced ostensibly though selection of treatments that explicitly facilitated sage-grouse movement between seasonal reproductive habitats and larger-scale PACs. Model output had high concordance with treatments implemented closer to the start of the study period, which indicated the “best” sites may have already been treated and subsequent treated sites might yield limited returns. Importantly, both studies also stress that the models are meant to support the decision-making process at the local scale, and not supplant local knowledge. Still, use of simulated changes in sage-grouse habitat selection following conifer removal provides a useful tool for budget-limited managers to avoid implementation in areas with low benefit to sage-grouse and high risk of disturbance from intensive treatment.
The aforementioned studies contribute to a proliferation of research that explicitly incorporate general R&R into planning and prioritization at the local-scale within the context (implicitly or explicitly) of sage-grouse habitat requirements and distribution at mid-scales (e.g., Knutson et al., 2014; Pyke et al., 2015; Chambers et al., 2017, 2019b; Barnard et al., 2019). However, few have modeled specifically quantified changes in sage-grouse habitat selection post-restoration as a function of predicted land cover responses to underlying general resilience. The aforementioned Ricca et al. (2018) study provides an example for local-scale fire restoration that builds on the conifer removal example through simulation of land cover recovery on pixel by pixel basis in a GIS given: (1) burn severity driving likely land cover change, (2) decisions to conduct passive or active restoration, (3) pre-burn land cover composition, fuel type, and underlying R&R class, and (4) uncertainty in resistance to annual grass invasion for moderate and low RR classes (Figure 8). For example, while active or passive restoration in high R&R yielded return to sagebrush or original land cover type, active restoration in low R&R could yield mixed sagebrush establishment under a resistant outcome but annual grass monocultures under a non-resistant outcome. Application of pre-fire RSF coefficients describing sage-grouse habitat selection responses to simulated post-fire landscapes under the different restored and/or resistant outcomes allows ranking of average sage-grouse habitat selection post-fire among sets of candidate wildfire scars examined, as with the conifer example. However, wildfire often immediately impacts thousands of acres of sagebrush. Hence, spatial heterogeneity in post-fire sage-grouse habitat selection can also be visualized though these types of decision-support tools to provide managers better identification of targeted areas for restoration across larger burned landscapes. These areas could include isolated patches of readily restorable habitat likely to improve connectivity within juxtaposed larger patches of less-recoverable annual grass, and areas where restoration could lessen risks of annual grass invasion on peripheries of more resilient and resilient patches, thereby increasing suitability across larger patch sizes. Moreover, relatively simple rules for land cover conversion in the decision tree can be expanded with parameters describing the efficacy of different treatment types and variation in sagebrush or herbaceous recovery processes relative to finer-scale variation in underlying general resilience depicted in more specialized mapping layers, and (as with the conifer tools) can be expanded to mid-to-broad scales using generalizable models informed by sage-grouse habitat selection parameters measured across multiple sites (see section: Pathways for Improving Decision-Support Metrics).
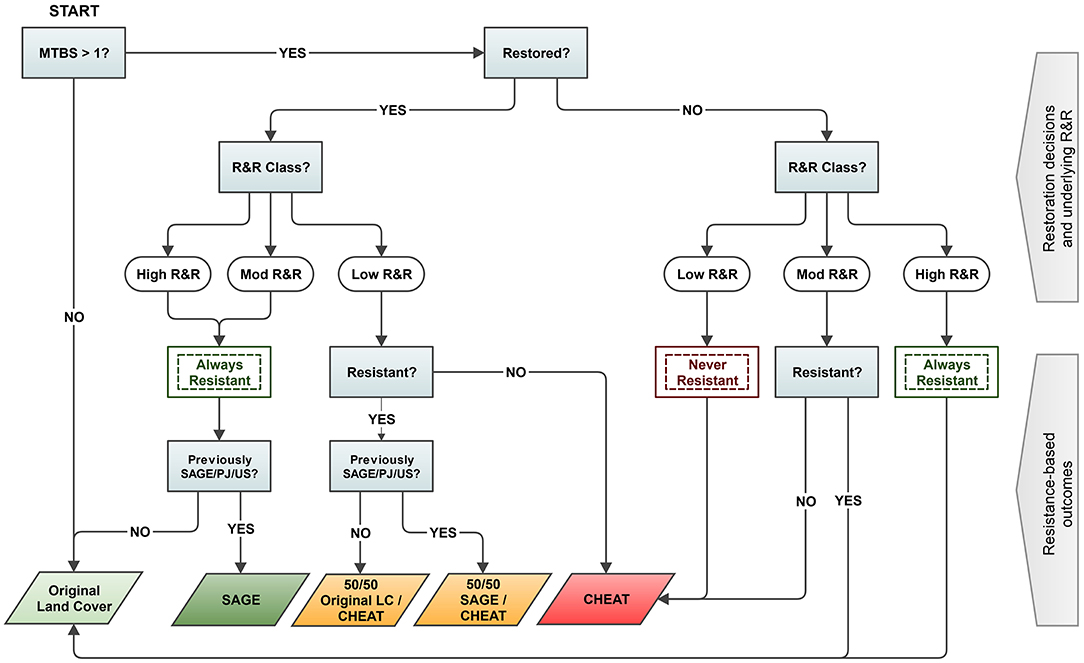
Figure 8. Diagram of the decision model used to simulate outcomes of restoration decisions on land cover change (900 m2) variation in burn-severity and resilience to disturbance and resistance to invasion (RandR) index classes. Local conditions may add uncertainty for resistance to cheatgrass invasion under some moderate and low RandR soil temperature and moisture regimes, so the model generates both Resistant and Not-Resistant outcomes in these cases following the decision to restore or not restore. Restored decisions assume active seeding or planting of sagebrush. Subsequent post-fire land cover surfaces can be used to model changes in sage-grouse habitat selection or suitability. Used with permission from Ricca et al. (2018).
Lastly, the above examples used mid-scale models to aid local decision support, but existing site-specific information on sage-grouse resource selection during critical life history periods prior to disturbance can also be leveraged opportunistically. The possibility for such data sets is increasingly likely, given the preponderance of sage-grouse studies employing intensive monitoring of marked sage-grouse to measure vital rates and spatial utilization across numerous sites in the fire-prone Great Basin, and more frequent and larger wildfires intersecting these sites. We provide an example of this scenario to help inform targeted restoration actions within the scar of a 1,277-km2 megafire in northeastern California (Rush Fire) that occurred in 2012. This example is part of a larger and collaborative on-going project evaluating sage-grouse spatial and demographic responses to restoration treatments in expansively burned landscapes, and forth-coming papers will describe overall project objectives and information regarding specific sagebrush planting design, seedling survival, and sage-grouse responses. General statistical methods for this example are described in the Supplementary Material Appendix I. In brief, an extensive dataset of sage-grouse nesting locations (Davis et al., 2014) informed an RSF describing pre-fire nesting habitat selection. Using GIS simulation approach described heretofore, we: (1) converted burned pixels to bare-ground, (2) recalculated habitat availability with appropriate moving windows immediately following the fire, (3) applied pre-fire RSF coefficients to the post-fire landscape, (4) subtracted the post- from pre-RSF to describe relative loss of nesting habitat (i.e., ΔRSFnesting), and (5) categorize the ΔRSFnesting by the 50th percentile to identify core nesting loss (Figures 8A–C). Intersecting core nesting loss with R&R (Figure 9D) then facilitates more surgical targeting of intensive restoration in areas of the greatest loss of nesting habitat along a gradient of elevation and R&R and juxtaposed to existing leks (Figures 8E–G). Planting sagebrush seedlings in heterogeneously dense patches designed to provide minimum nesting cover within ~3–4 years incorporates a resource island approach to restoration (Hulvey et al., 2017), which accounts for sage-grouse site fidelity to nesting sites despite disturbance, and can subsequently help ameliorate asynchronies in engineering resilience between recovering sage-grouse and sagebrush populations post-fire.
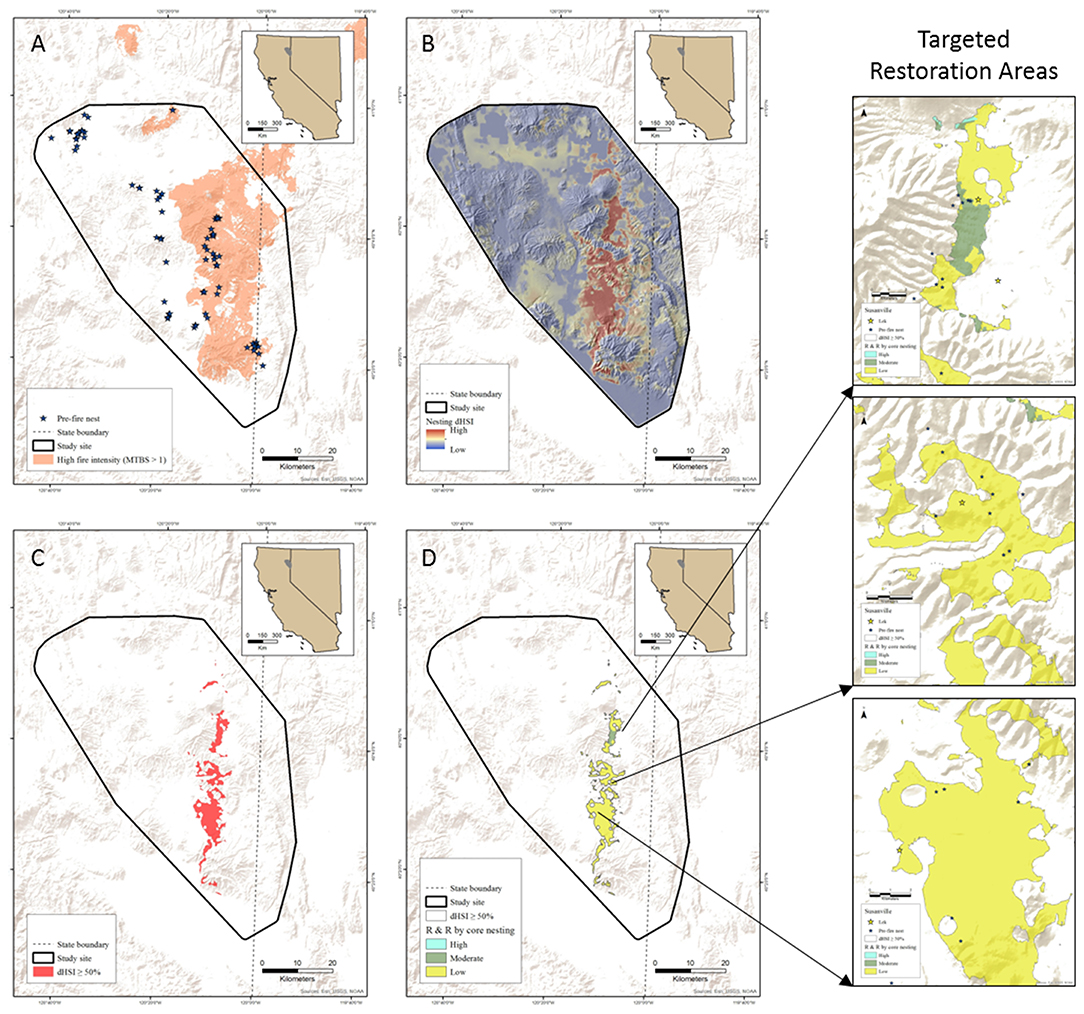
Figure 9. Example of identifying areas for targeted restoration of sage-grouse nesting habitat affected by the Rush Fire in northeastern California. Pre-fire sage-grouse nest locations (A) inform a resource selection function of habitat loss post-fire (B), which is categorized by the 50th percentile representing core loss (C) and intersected with RandR (D).
Pathways For Improving Decision Support Metrics
The examples help bridge gaps in linking resilience concepts based on plant and soil processes driving ecosystem productivity with responses of higher trophic level and indicator species such as sage-grouse. A key to this process is identification of spatial relationships describing sage-grouse distribution and habitat selection with predicted sage-grouse and sagebrush ecosystem responses to disturbance and subsequent restoration efforts. Mid-scale examples focus on prioritization of active and passive actions using a geospatial overlay approach derived from the Framework. Telemetry-based models of habitat selection and high-resolution mapping of conifer canopy cover could be expanded to the broad-scale as soon as rangewide companion models are derived. Changes in land cover composition given underlying R&R using local scale models allow further quantification of spatial heterogeneity in improvement in sage-grouse habitat selection following disturbance and restoration, and those reorganized process can be scaled hierarchically to mid-and-broad scales (Perring et al., 2015) using generalizable models informed by datasets spanning multiple sites and years. Identification of temporal mismatches in engineering resilience between sage-grouse and sagebrush population dynamics impacts provides an explanation for the overall processes driving negative sage-grouse population growth with cumulative impacts of wildfire. Nevertheless, the decision support tools we described can be improved greatly through more explicit quantification of impacts on sage-grouse fitness and movement connectivity following disturbance and restoration under different scenarios of changing land cover composition, and by adding more complexity to sagebrush recovery models to better reflect spatial heterogeneity in feedbacks driving resilience.
Incorporating Sage-Grouse Fitness Consequences
Analyses that quantify disproportionate use or avoidance (e.g., resource selection functions) are often used to infer suitability resource configurations for meeting life history needs. Yet, the true measure of suitability relates to differential fitness in terms increased survival and reproduction leading to population stability or growth (Hirzel and Le Lay, 2008; Gaillard et al., 2010). Inferences of suitability from selection can be confounded by existence of density-dependent source-sink dynamics (Matthiopoulos et al., 2015) and the aforementioned ecological traps, which are known to exist across sage-grouse populations (Aldridge and Boyce, 2007; Coates et al., 2017b; Heinrichs et al., 2018) but can be difficult to quantify due to modeling complexity and data limitations across broad spatiotemporal scales and finer grain. Recent advances in hierarchical modeling frameworks and computing power can help facilitate a shift from resource selection- to fitness-based metrics for use as ecological currency in decision support tools.
For example, significant advances have been made in spatially explicit estimation of population change (i.e., lambda) derived from demographic matrix or integrated population models that share information across multiple datasets and account for observation error (Chandler et al., 2018). Subsequent output can provide more refined geospatial overlays depicting areas with predicted population growth under both current and disturbance-induced conditions parametrized as multi-scale land cover covariates, which help identify specific habitats whose loss correlates with reductions in population growth and provide another measure for prioritization of prevention, suppression, and restoration efforts (e.g., Figures 3–5). Hierarchical Bayesian models also allow ready linkage of posterior parameter distributions describing resource selection during specific life-stages with those describing concomitant survival probabilities (Coates et al., 2017b). Such approaches were used recently in a generalizable mid-to-broad scale model that depicted source-sink dynamics in relation to habitat features and underlying coarse-scale R&R conditions influencing sage-grouse nest selection and survival across much of the Great Basin (O'Neil et al. unpublished manuscript). That study also determined that underlying R&R conditions mediated sage-grouse functional responses to habitat features (e.g., stronger selection for sagebrush in areas with low R&R), yet nest survival increased concomitantly with R&R. Similarly, individual-based models that simulate how demographic outcomes across an individual's life cycle are modulated by interactions with changing landscape features have been constructed to map source-sink dynamics on the northern periphery of sage-grouse range (Heinrichs et al., 2018). Spatially explicit depictions of how sage-grouse gene-flow at mid-to-broad scales is constricted once reductions in habitat suitability surpass thresholds indexing impermeability to movement that further inform estimates of meta-population persistence (Fedy et al., 2016; Row et al., 2018). Spatially explicit layers from such studies fit nicely in geospatial prioritization overlays, and changes in fitness landscapes following land cover change from disturbance or restoration as mediated by R&R can be estimated directly in a GIS using model-derived parameters. Moreover, fitness-based measures help better identify thresholds of resource loss (Standish et al., 2014; Chambers et al., 2019a) that tip populations from growth or stability to decline and provide a quantification of energy needed to surpass ecological resilience into undesired states.
Improving Estimates of Sagebrush Engineering and Spatial Resilience
The examples we described integrate sage-grouse metrics with coarse, 3-level indices of R&R (Maestas et al., 2016), which represented the first generation of spatially explicit estimates of general R&R in the original Framework (Chambers et al., 2017). These indices represent an aggregation of much finer soil temperature and subclasses that provided a highly tractable approach for managers faced with decision making at broad to local scales from spatial overlays and corresponding 3 x 3 decision matrices. Accordingly, they provided a useable and novel means for modeling sage-grouse population dynamics (Coates et al., 2016a) and habitat selection (Coates et al., 2016b; Ricca et al., 2018, other examples heretofore) as a function of resource availability following disturbance as mediated by underlying R&R. However, a strength of the Framework and subsequent approaches that follow is the ability to readily incorporate newly available information. Such information is now proliferating in the literature, and we provide general pathways for incorporation of new models of sagebrush population dynamics, state-transition, and general resilience to produce more refined predictors of engineering and spatial resilience.
First, coupling sage-grouse population growth models (e.g., Coates et al., 2016a) with better parameterized models of sagebrush population growth and corresponding estimates of cumulative burn area (CBA) should help ameliorate (or at least better identify and subsequently prioritize) mismatches in engineering resilience across spatial scales. For example, strong transient dynamics (i.e., where short term population trends following disturbance are decoupled from those resulting in undisturbed long-term trends) have been identified recently across seeded post-fire sagebrush populations across the western U.S., which arise due to altered population size structure post-disturbance and reduced survival and fecundity of seedlings compared to more robust and established plants (Shriver et al., 2019). The Shriver et al. (2019) study highlighted that while establishment is promoted during years of favorable wet and cool overwinter conditions, transient dynamics are often difficult to overcome due to low sagebrush density or outright failure in sites with predicted long-term sagebrush stability. Similarly, Requena-Mullor et al. (2019) demonstrated stronger effects of past local fire history (in terms of number and occurrence) compared to regional climate on big sagebrush occurrence and cover, while restoration only impacted occurrence and cover across the Great Basin. These results also further highlight the importance of conducting targeted restoration in areas with high reproductive value to breeding sage-grouse to help overcome differences in sage-grouse and sagebrush engineering resilience (Figure 1), such as the example we describe for the Rush Fire restoration (Figure 9). Using techniques such as seedling-based treatments that help bolster immediate survival probability and subsequently help overcome transient dynamics from seeded treatments, particularly if replicated across densely-planted patches that account for high first year seedling mortality (Brabec et al., 2015) could still provide cover for nesting sage-grouse in a short amount of time. Such efforts require substantial time and effort, which is another reason for a highly surgical approach.
Second, parameters from studies that explicitly model variation in sagebrush recovery processes as a function of underlying R&R can better inform predictions from state-transition models (e.g., Briske et al., 2008; Stringham et al., 2016; Chambers et al., 2017) and subsequent sage-grouse response. Similar to the studies described above, additional meta-analyses of space for time studies describing sagebrush recovery processes (e.g., Knutson et al., 2014; Barnard et al., 2019) following restoration (Pilliod and Welty, 2013; Pilliod et al., 2017b) in the context of spatially explicit R&R layers at coarse to fine scales (e.g., soil moisture and temperature sub-classes) would be especially useful; as would back-in-time approaches (Shi et al., 2017) that leverage extensive time series of archived satellite data (e.g., Landsat) across expansive extents to classify changes in land cover at relatively high resolution (e.g., percentages of functional plant types with 900 m2 pixels) (Xian et al., 2015) and then relate back to R&R in a similar fashion. For the latter case, Monroe et al. (2020) recently utilized a back-in-time approach to quantify factors influencing sagebrush recovery on reclaimed well-pads in Wyoming, and found that dynamic variables such as annual precipitation and temperature modified annual rates of change in cover (e.g., engineering resilience) based on more static state-variables such as soil type and topographic position describing general resilience. Specifically, growth rates increased more strongly following warm and wet conditions in higher elevation sites but declined with warmer conditions in lower elevation sites. Dynamic patterns in precipitation can also have strong, differential impacts on annual availability and drought resiliency of mesic resources across elevational and mid-scale ecoregional gradients (Donnelly et al., 2018). Models such as these that quantify modifications of static predictions of sagebrush engineering resilience by inter-annual variation in precipitation and temperature can further help parameterize decision-support tools for sage-grouse given that sage-grouse populations respond positively to pulses of above average precipitation at local- (Blomberg et al., 2012) and mid-scales (Coates et al., 2016a; Donnelly et al., 2018).
Third, soil-based geospatial layers describing layers general resilience can be broken down into finer levels of organization for subsequent use in predictive models of sagebrush recovery and state-transition, and then substituted in sage-grouse geospatial overlay or simulation analyses. For example, the aggregated 3-class R&R index of Maestas et al. (2016) can, and has been, deconstructed into finer subclasses of soil moisture and temperature regimes, with companion state-transition models nested within ecoregion and major land use area type across the eastern portion of sage-grouse range (Chambers et al., 2016). A similar framework has been developed for much of the Great Basin that aggregates local sites with different ecological potential, as governed by soil and climate conditions, into more manageable yet still fine-scale groupings based on shared predicted responses to disturbance and associated state-transitions also nested within major land use areas (i.e., DRGs, Stringham et al., 2016). Newly developed mid-scale spatially explicit models that leverage multi-decadal measures of plant-greenness with biophysical covariates (e.g., topographic position, soil organic matter, and available water capacity) to map disproportionate deviation of current vegetation composition and structure from estimated site-potential along elevational and disturbance gradients across the Great Basin represent a very powerful new tool (Rigge et al., 2019). Development of spatially explicit models depicting variation in soil macro and microbial biota, which also influence resistance to annual grass invasion (Belnap and Phillips, 2001; Bansal and Sheley, 2016), would allow novel incorporation of rather under-studied yet important soil process. Collectively, layers such as this help downscale coarser R&R predictions of potential community state-transitions following disturbance given finer-scale edaphic, topographic, and climatic conditions, and are better informed by parameters derived from empirical models described above. However, spatially explicit modeling of state-transitions is still in a nascent stage, largely due to difficulty in parameterizing complex processes and common reliance on information synthesized from the literature or expert opinion. While still very useful, practitioners who use these tools need to be cognizant that model output is determined largely by user-defined deterministic rules and requires independent validation (Requena-Mullor et al., 2019; Chambers et al., 2019b). Importantly, the conservation planning tools simulating changes in land cover given underlying R&R that we described, even with greater modeled complexity, also fall under this same caveat.
Conclusion
Model-based efforts toward operationalizing resilience in sagebrush ecosystems show significant impact. For example, a Web of Science search (model* and resilience and restoration) listed Briske et al. (2008) who first described incorporation of resilience into rangeland state-transition models, and Chambers et al. (2014) as described above, as the 9 and 10th most cited papers, respectively. Moreover, a key benefit of this impact, and subsequent unprecedented efforts toward the conservation of the imperiled sagebrush biome over the past decade, has been an increasing integration of foundational and novel ideas from the fields of ecosystem restoration, rangeland, and wildlife ecology. Here, we provided multiple examples using geospatial overlays and simulation approaches that demonstrated how multi-scale linkages of underlying general resilience with “sage-grouse centric” measures of population performance and resource availability. Results yield highly tractable decision-support tools for real-world managers that help increase interconnected ecological, engineering, and spatial resilience for both sage-grouse populations and sagebrush ecosystems. We note that while our examples focused on operationalizing resilience to conifer and wildfire, similar approaches could be applied to other disturbances such as energy and agricultural development and help guide grazing regimes for livestock and free-ranging equids given appropriate parameterization. We also recognize that the multiple and somewhat independent decision support tools presented could be confusing for managers in need of more comprehensive tools housed in a single, “one-stop-shopping” type of framework. Tools that differentially weight different desired outcomes through structured decision models (e.g., Martin et al., 2009) made spatially-explicit would help bridge this gap for managers. Lastly, we stress the need for structured long-term monitoring of focal species responses to validate predicted outcomes of restoration from these approaches (Suding, 2011). Sage-grouse are also a highly useful species to link plant and soil with higher trophic responses, yet the concepts and tools we described can be applied using other indicator species or assemblages of species, including those with different life-history strategies and resource needs that might not be covered always under the umbrella of sage-grouse (Carlisle et al., 2018).
Data Availability Statement
Geospatial layers are available for download at the USGS ScienceBase website. Prioritization examples for wildfire and conifer management (Figures 3, 4, 6) are available at https://doi.org/10.5066/P960W8MD. Additional layers are available at https://doi.org/10.5066/F7G15ZRN, https://doi.org/10.5066/F7K35RRS, and https://doi.org/10.5066/F7TT4Q5S.
Ethics Statement
Sage-grouse marking efforts for the authors' field studies that generated data, in part, for this manuscript were vetted and approved by the USGS-Western Ecological Animal Use and Care Committee (USGS_ACUC-002).
Author Contributions
PC and MR conceived the ideas and models described within that integrate sage-grouse and sagebrush R&R metrics. MR and PC wrote the manuscript and gave final approval for publication.
Funding
This work was provided by U.S. Geological Survey, Bureau of Land Management, U.S. Fish and Wildlife Service, U.S. Forest Service, Nevada Department of Wildlife, California Department of Fish and Wildlife, and Ormat Technologies.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Many of the examples presented in this manuscript were synthesized initially during a symposium on the Framework held at the 2018 annual meeting of the Society of Rangeland Management. We thank J. Chambers for this opportunity to present our work in this special edition. We are extremely appreciative for expert statistical and GIS support from B. Prochazka, B. Brussee, M. Chenaille, B. Gustafson, T. Kroger, S. O'Neil, C. Roth, and E. Sanchez-Chopitea. The Rush Fire restoration example is part of larger project conceived and executed in partnership with C. Aldridge, D. Davis, S. Hanser, J. Heinrichs, and D. Pyke. We thank countless field technicians who collected data that informed sage-grouse models, and the federal, state, and private clients who supported many of the studies described in this paper, including (but not limited to) U.S. Geological Survey, Bureau of Land Management, U.S. Fish and Wildlife Service, U.S. Forest Service, Nevada Department of Wildlife, California Department of Fish and Wildlife, and Ormat Technologies. We appreciate constructive manuscript reviews from J. Atkinson, T. Kimball, K. Miles, J. Severson, J. Vogt, and referees.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00493/full#supplementary-material
References
Aldridge, C. L., and Boyce, M. S. (2007). Linking occurrence and fitness to persistence: habitat-based approach for endangered Greater Sage-Grouse. Ecol. Appl. 17, 508–526. doi: 10.1890/05-1871
Andelman, S. J., and Fagan, W. F. (2000). Umbrellas and flagships: efficient conservation surrogates or expensive mistakes? Proc. Natl. Acad. Sci. U.S.A. 97, 5954–5959. doi: 10.1073/pnas.100126797
Angeler, D. G., and Allen, C. R. (2016). Quantifying resilience. J. Appl. Ecol. 53, 617–624. doi: 10.1111/1365-2664.12649
Arkle, R. S., Pilliod, D. S., Hanser, S. E., Brooks, M. L., Chambers, J. C., Grace, J. B., et al. (2014). Quantifying restoration effectiveness using multi-scale habitat models: implications for sage-grouse in the Great Basin. Ecosphere 5:art31. doi: 10.1890/ES13-00278.1
Balch, J. K., Bradley, B. A., D'Antonio, C. M., and Gómez-Dans, J. (2013). Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Glob. Change Biol. 19, 173–183. doi: 10.1111/gcb.12046
Bansal, S., and Sheley, R. L. (2016). Annual grass invasion in sagebrush steppe: the relative importance of climate, soil properties and biotic interactions. Oecologia 181, 543–557. doi: 10.1007/s00442-016-3583-8
Barnard, D. M., Germino, M. J., Arkle, R. S., Bradford, J. B., Duniway, M. C., Pilliod, D. S., et al. (2019). Soil characteristics are associated with gradients of big sagebrush canopy structure after disturbance. Ecosphere 10:e02780. doi: 10.1002/ecs2.2780
Baruch-Mordo, S., Evans, J. S., Severson, J. P., Naugle, D. E., Maestas, J. D., Kiesecker, J. M., et al. (2013). Saving sage-grouse from the trees: a proactive solution to reducing a key threat to a candidate species. Biol. Conserv. 167, 233–241. doi: 10.1016/j.biocon.2013.08.017
Battin, J. (2004). When good animals love bad habitats: ecological traps and the conservation of animal populations. Conserv. Biol. 18, 1482–1491. doi: 10.1111/j.1523-1739.2004.00417.x
Beisner, B. E., Haydon, D. T., and Cuddington, K. (2003). Alternative stable states in ecology. Front. Ecol. Environ. 1, 376–382. doi: 10.1890/1540-9295(2003)001[0376:ASSIE]2.0.CO;2
Belnap, J., and Phillips, S. L. (2001). Soil biota in an ungrazed grassland: response to annual grass (Bromus tectorum) invasion. Ecol. Appl. 11, 1261–1275. doi: 10.1890/1051-0761(2001)011[1261:SBIAUG]2.0.CO;2
Bender, E. A., Case, T. J., and Gilpin, M. E. (1984). Perturbation experiments in community ecology: theory and practice. Ecology 65, 1–13. doi: 10.2307/1939452
Bestelmeyer, B. T., Ellison, A. M., Fraser, W. R., Gorman, K. B., Holbrook, S. J., Laney, C. M., et al. (2011). Analysis of abrupt transitions in ecological systems. Ecosphere 2:129. doi: 10.1890/ES11-00216.1
Blomberg, E. J., Sedinger, J. S., Atamian, M. T., and Nonne, D. V. (2012). Characteristics of climate and landscape disturbance influence the dynamics of greater sage-grouse populations. Ecosphere 3:art55. doi: 10.1890/ES11-00304.1
Boyte, S. P., Wylie, B. K., and Major, D. J. (2019). Validating a time series of annual grass percent cover in the sagebrush ecosystem. Rangeland Ecol. Manage. 72, 347–359. doi: 10.1016/j.rama.2018.09.004
Brabec, M. M., Germino, M. J., Shinneman, D. J., Pilliod, D. S., McIlroy, S. K., and Arkle, R. S. (2015). Challenges of establishing big sagebrush (Artemisia tridentata) in rangeland restoration: effects of herbicide, mowing, whole-community seeding, and sagebrush seed sources. Rangeland Ecol. Manage. 68, 432–435. doi: 10.1016/j.rama.2015.07.001
Bradley, B. A., Curtis, C. A., Fusco, E. J., Abatzoglou, J. T., Balch, J. K., Dadashi, S., et al. (2017). Cheatgrass (Bromus tectorum) distribution in the intermountain Western United States and its relationship to fire frequency, seasonality, and ignitions. Biol. Invas. 20, 1493–1506. doi: 10.1007/s10530-017-1641-8
Briske, D. D., Bestelmeyer, B. T., Stringham, T. K., and Shaver, P. L. (2008). Recommendations for development of resilience-based state-and-transition models. Rangeland Ecol. Manage. 61, 359–367. doi: 10.2111/07-051.1
Brooks, M. L., Matchett, J. R., Shinneman, D. J., and Coates, P. S. (2015). Fire Patterns in the Range of the Greater Sage-Grouse, 1984-2013 — Implications for Conservation and Management. U.S. Geological Survey Open-File Report 2015–1167. doi: 10.3133/ofr20151167
Bybee, J., Roundy, B. A., Young, K. R., Hulet, A., Roundy, D. B., Crook, L., et al. (2016). Vegetation response to piñon and juniper tree shredding. Rangeland Ecol. Manage. 69, 224–234. doi: 10.1016/j.rama.2016.01.007
Carlisle, J. D., Keinath, D. A., Albeke, S. E., and Chalfoun, A. D. (2018). Identifying holes in the greater sage-grouse conservation umbrella. J. Wildlife Manage. 82, 948–957. doi: 10.1002/jwmg.21460
Carpenter, S. R., Mooney, H. A., Agard, J., Capistrano, D., Defries, R. S., Díaz, S., et al. (2009). Science for managing ecosystem services: beyond the Millennium Ecosystem Assessment. Proc. Natl. Acad. Sci. U.S.A. 106, 1305–1312. doi: 10.1073/pnas.0808772106
Chambers, J. C., Allen, C. R., and Cushman, S. A. (2019a). Operationalizing ecological resilience concepts for managing species and ecosystems at risk. Front. Ecol. Evol. 7:241. doi: 10.3389/fevo.2019.00241
Chambers, J. C., Beck, J. L., Bradford, J. B., Bybee, J., Campbell, S., Carlson, J., et al. (2017). Science Framework for Conservation and Restoration of the Sagebrush Biome: Linking the Department of the Interior's Integrated Rangeland Fire Management Strategy to Long-term Strategic Conservation Actions. Gen. Tech. Rep. RMRS-GTR-360. U.S Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, CO, 213.
Chambers, J. C., Beck, J. L., Campbell, S., Carlson, J., Christensen, T. J., Clause, K. J., et al. (2016). Using Resilience and Resistance Concepts to Manage Threats to Sagebrush Ecosystems, Gunnison Sage-Grouse, and Greater Sage-Grouse in Their Eastern Range: A Strategic Multi-Scale Approach. Gen. Tech. Rep. RMRS-GTR-356. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, CO, 143.
Chambers, J. C., Bradley, B. A., Brown, C. S., D'Antonio, C., Germino, M. J., Grace, J. B., et al. (2014). Resilience to stress and disturbance, and resistance to Bromus tectorum L. invasion in cold desert shrublands of Western North America. Ecosystems 17, 360–375. doi: 10.1007/s10021-013-9725-5
Chambers, J. C., Brooks, M. L., Germino, M. J., Maestas, J. D., Board, D. I., Jones, M. O., et al. (2019b). Operationalizing resilience and resistance concepts to address invasive grass-fire cycles. Front. Ecol. Evol. 7:185. doi: 10.3389/fevo.2019.00185
Chambers, J. C., Roundy, B. A., Blank, R. R., Meyer, S. E., and Whittaker, A. (2007). What makes Great Basin sagebrush ecosystems invasible by Bromus tectorum? Ecol. Monogr. 77, 117–145. doi: 10.1890/05-1991
Chandler, R. B., Hepinstall-Cymerman, J., Merker, S., Abernathy-Conners, H., and Cooper, R. J. (2018). Characterizing spatio-temporal variation in survival and recruitment with integrated population models. Auk 135, 409–426. doi: 10.1642/AUK-17-181.1
Chapin, F. S., Sommerkorn, M., Robards, M. D., and Hillmer-Pegram, K. (2015). Ecosystem stewardship: a resilience framework for arctic conservation. Glob. Environ. Change 34, 207–217. doi: 10.1016/j.gloenvcha.2015.07.003
Coates, P. S., Brussee, B. E., Ricca, M. A., Dudko, J. E., Prochazka, B. G., Espinosa, S. P., et al. (2017a). Greater Sage-Grouse (Centrocercus urophasianus) Nesting and Brood-Rearing Microhabitat in Nevada and California—Spatial Variation in Selection and Survival Patterns. Report 2017-1087, Reston, VA. doi: 10.3133/ofr20171087
Coates, P. S., Brussee, B. E., Ricca, M. A., Severson, J. P., Casazza, M. L., Gustafson, K. B., et al. (2019). Spatially explicit models of seasonal habitat for greater sage-grouse at broad spatial scales: informing areas for management in Nevada and northeastern California. Ecol. Evol. doi: 10.1002/ece3.5842. [Epub ahead of print].
Coates, P. S., Casazza, M. L., Blomberg, E. J., Gardner, S. C., Espinosa, S. P., Yee, J. L., et al. (2013). Evaluating greater sage-grouse seasonal space use relative to leks: Implications for surface use designations in sagebrush ecosystems. J. Wildlife Manage. 77, 1598–1609. doi: 10.1002/jwmg.618
Coates, P. S., Casazza, M. L., Ricca, M. A., Brussee, B. E., Blomberg, E. J., Gustafson, K. B., et al. (2016b). Integrating spatially explicit indices of abundance and habitat quality: an applied example for greater sage-grouse management. J. Appl. Ecol. 53, 83–95. doi: 10.1111/1365-2664.12558
Coates, P. S., Prochazka, B. G., Ricca, M. A., Gustafson, K. B., Ziegler, P., and Casazza, M. L. (2017b). Pinyon and juniper encroachment into sagebrush ecosystems impacts distribution and survival of greater sage-grouse. Rangeland Ecol. Manage. 70, 25–38. doi: 10.1016/j.rama.2016.09.001
Coates, P. S., Ricca, M. A., Prochazka, B. G., Brooks, M. L., Doherty, K. E., Kroger, T. E., et al. (2016a). Wildfire, climate, and invasive grass interactions negatively impact an indicator species by reshaping sagebrush ecosystems. Proc. Natl. Acad. Sci. U.S.A. 113, 12745–12750. doi: 10.1073/pnas.1606898113
Crist, M. R., Chambers, J. C., Phillips, S. L., Prentice, K. L., and Wiechman, L. A. (2019). Science Framework for Conservation and Restoration of the Sagebrush Biome: Linking the Department of the Interior's Integrated Rangeland Fire Management Strategy to Long-Term Strategic Conservation Actions. Part 2. Management applications. Gen. Tech. Rep. RMRS-GTR-389. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, CO, 237. doi: 10.2737/RMRS-GTR-389
Davies, K. W., Boyd, C. S., Beck, J. L., Bates, J. D., Svejcar, T. J., and Gregg, M. A. (2011). Saving the sagebrush sea: an ecosystem conservation plan for big sagebrush plant communities. Biol. Conserv. 144, 2573–2584. doi: 10.1016/j.biocon.2011.07.016
Davis, D. M., Reese, K. P., and Gardner, S. C. (2014). Demography, reproductive ecology, and variation in survival of greater sage-grouse in northeastern California. J. Wildlife Manage. 78, 1343–1355. doi: 10.1002/jwmg.797
Department of Interior (2015). Secretary of the Interior Order No. 3336 - Rangeland Fire Prevention, Management, and Restoration. Available online at: https://www.nifc.gov/fireandsagegrouse/docs/SO_Rangeland.pdf.
Doherty, K. E., Evans, J. S., Coates, P. S., Juliusson, L. M., and Fedy, B. C. (2016). Importance of regional variation in conservation planning: a rangewide example of the Greater Sage-Grouse. Ecosphere 7:e01462. doi: 10.1002/ecs2.1462
Doherty, K. E., Hennig, J. D., Dinkins, J. B., Griffin, K. A., Cook, A. A., Maestas, J. D., et al. (2018). Understanding biological effectiveness before scaling up range-wide restoration investments for Gunnison sage-grouse. Ecosphere 9:e02144. doi: 10.1002/ecs2.2144
Donnelly, J. P., Allred, B. W., Perret, D., Silverman, N. L., Tack, J. D., Dreitz, V. J., et al. (2018). Seasonal drought in North America's sagebrush biome structures dynamic mesic resources for sage-grouse. Ecol. Evol. 8:12492–12505. doi: 10.1002/ece3.4614
Donnelly, J. P., Naugle, D. E., Hagen, C. A., and Maestas, J. D. (2016). Public lands and private waters: scarce mesic resources structure land tenure and sage-grouse distributions. Ecosphere 7:e01208. doi: 10.1002/ecs2.1208
Donnelly, J. P., Tack, J. D., Doherty, K. E., Naugle, D. E., Allred, B. W., and Dreitz, V. J. (2017). Extending conifer removal and landscape protection strategies from sage-grouse to songbirds, a range-wide assessment. Rangeland Ecol. Manage. 70, 95–105. doi: 10.1016/j.rama.2016.10.009
Eidenshink, J., Schwind, B., Brewer, K., Zhu, Z. L., Quayle, B., and Howard, S. (2007). A project for monitoring trends in burn severity. Fire Ecol. 3, 3–22. doi: 10.4996/fireecology.0301003
Ernst-Brock, C., Turner, L., Tausch, R. J., and Leger, E. A. (2019). Long-term vegetation responses to pinyon-juniper woodland reduction treatments in Nevada, USA. J. Environ. Manage. 242, 315–326. doi: 10.1016/j.jenvman.2019.04.053
Eviner, V. T., and Hawkes, C. V. (2008). Embracing variability in the application of plant–soil interactions to the restoration of communities and ecosystems. Restor. Ecol. 16, 713–729. doi: 10.1111/j.1526-100X.2008.00482.x
Falkowski, M. J., Evans, J. S., Naugle, D. E., Hagen, C. A., Carleton, S. A., Maestas, J. D., et al. (2017). Mapping tree canopy cover in support of proactive prairie grouse conservation in western North America. Rangeland Ecol. Manage. 70, 15–24. doi: 10.1016/j.rama.2016.08.002
Fedy, B. C., Row, J. R., and Oyler-McCance, S. J. (2016). Integration of genetic and demographic data to assess population risk in a continuously distributed species. Conserv. Genet. 18, 89–104. doi: 10.1007/s10592-016-0885-7
Folke, C., Carpenter, S., Walker, B., Scheffer, M., Elmqvist, T., Gunderson, L., et al. (2004). Regime shifts, resilience, and biodiversity in ecosystem management. Annu. Rev. Ecol. Evol. Syst. 35, 557–581. doi: 10.1146/annurev.ecolsys.35.021103.105711
Foster, L., Dugger, K. M., Hagen, C., and Budeau, D. A. (2019). Greater sage-grouse vital rates after wildfire. J. Wildlife Manage. 83, 121–134. doi: 10.1002/jwmg.21573
Gaillard, J.-M., Hebblewhite, M., Loison, A., Fuller, M., Powell, R., Basille, M., et al. (2010). Habitat–performance relationships: finding the right metric at a givenspatial scale. Philos. Trans. R. Soc. B Biol. Sci. 365, 2255–2265. doi: 10.1098/rstb.2010.0085
Germino, M. J., Barnard, D. M., Davidson, B. E., Arkle, R. S., Pilliod, D. S., Fisk, M. R., et al. (2018). Thresholds and hotspots for shrub restoration following a heterogeneous megafire. Landsc. Ecol. 33, 1177–1194. doi: 10.1007/s10980-018-0662-8
Germino, M. J., Belnap, J., Stark, J. M., Allen, E. B., and Rau, B. M. (2016). “Ecosystem impacts of exotic annual invaders in the genus Bromus,” in Exotic Brome-Grasses in Arid and Semiarid Ecosystems of the Western US. Eds M. J. Germino, J. C. Chambers, and C. Brown (Berlin: Springer), 61–95. doi: 10.1007/978-3-319-24930-8_3
Gunderson, L. H. (2000). Ecological resilience - In theory and application. Annu. Rev. Ecol. Syst. 31, 425–439. doi: 10.1146/annurev.ecolsys.31.1.425
Gustafson, K. B., Coates, P. S., Roth, C. L., Chenaille, M. P., Ricca, M. A., Sanchez-Chopitea, E., et al. (2017). Using object-based image analysis to conduct high- resolution conifer extraction at regional spatial scales. Int. J. Appl. Earth Observ. Geoinform. 73, 148–155. doi: 10.1016/j.jag.2018.06.002
Hanser, S. E., and Knick, S. T. (2011). “Greater sage-grouse as an umbrella species for shrubland birds: a multi-scale assessment,” in Greater Sage-Grouse: Ecology and Conservation of a Landscape Species and Its Habitats, eds S. T. Knick and J. W. Connelly (Berkeley, CA: University of California Press), 475–487. doi: 10.1525/california/9780520267114.003.0020
Heinrichs, J. A., Aldridge, C. L., Gummer, D. L., Monroe, A. P., and Schumaker, N. H. (2018). Prioritizing actions for the recovery of endangered species: emergent insights from Greater Sage-grouse simulation modeling. Biol. Conserv. 218, 134–143. doi: 10.1016/j.biocon.2017.11.022
Hirzel, A. H., and Le Lay, G. (2008). Habitat suitability modelling and niche theory. J. Appl. Ecol. 45, 1372–1381. doi: 10.1111/j.1365-2664.2008.01524.x
Hobbs, R. J., Higgs, E., and Harris, J. A. (2009). Novel ecosystems: implications for conservation and restoration. Trends Ecol. Evol. 24, 599–605. doi: 10.1016/j.tree.2009.05.012
Holling, C. S. (1973). Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 4, 1–23. doi: 10.1146/annurev.es.04.110173.000245
Hulvey, K. B., Leger, E. A., Porensky, L. M., Roche, L. M., Veblen, K. E., Fund, A., et al. (2017). Restoration islands: a tool for efficiently restoring dryland ecosystems? Restor. Ecol. 25, S124–S134. doi: 10.1111/rec.12614
Knick, S. T., Hanser, S. E., and Preston, K. L. (2013). Modeling ecological minimum requirements for distribution of greater sage-grouse leks: implications for population connectivity across their western range U.S.A. Ecol. Evol. 3, 1539–1551. doi: 10.1002/ece3.557
Knutson, K. C., Pyke, D. A., Wirth, T. A., Arkle, R. S., Pilliod, D. S., Brooks, M. L., et al. (2014). Long-term effects of seeding after wildfire on vegetation in Great Basin shrubland ecosystems. J. Appl. Ecol. 5, 1414–1424. doi: 10.1111/1365-2664.12309
Kormos, P. R., Marks, D., Pierson, F. B., Williams, C. J., Hardegree, S. P., Havens, S., et al. (2017). Ecosystem water availability in juniper versus sagebrush snow-dominated rangelands. Rangeland Ecol. Manage. 70, 116–128. doi: 10.1016/j.rama.2016.05.003
Maestas, J. D., Campbell, S. B., Chambers, J. C., Pellant, M., and Miller, R. F. (2016). Tapping soil survey information for rapid assessment of sagebrush ecosystem resilience and resistance. Rangelands 38, 120–128. doi: 10.1016/j.rala.2016.02.002
Manier, D. J., Bowen, Z. H., Brooks, M. L., Casazza, M. L., Coates, P. S., Deibert, P. A., et al. (2014). Conservation Buffer Distance Estimates for Greater Sage-Grouse—A Review. U.S. Geological Survey Open-File Report 2014–1239. doi: 10.3133/ofr20141239
Martin, J., Runge, M. C., Nichols, J. D., Lubow, B. C., and Kendall, W. L. (2009). Structured decision making as a conceptual framework to identify thresholds for conservation and management. Ecol. Appl. 19, 1079–1090. doi: 10.1890/08-0255.1
Matthiopoulos, J., Fieberg, J., Aarts, G., Beyer, H. L., Morales, J. M., and Haydon, D. T. (2015). Establishing the link between habitat selection and animal population dynamics. Ecol. Monogr. 85, 413–436. doi: 10.1890/14-2244.1
Miller, J. R., and Hobbs, R. J. (2007). Habitat restoration—Do we know what we're doing? Restor. Ecol. 15, 382–390. doi: 10.1111/j.1526-100X.2007.00234.x
Miller, R. F., Bates, J. D., Svejcar, T. J., Pierson, F. B., and Eddleman, L. E. (2005). Biology, Ecology, and Management of Western Juniper. Technical Bulletin 152. Oregon State University Agricultural Experiment Station. Available online at: http://juniper.oregonstate.edu/bibliography/documents/phpQ65pOk_tb152.pdf.
Monroe, A. P., Aldridge, C. L., O'Donnell, M. S., Manier, D. J., Homer, C. G., and Anderson, P. J. (2020). Using remote sensing products to predict recovery of vegetation across space and time following energy development. Ecol. Indicat. 110:105872. doi: 10.1016/j.ecolind.2019.105872
Palmer, M. A., Ambrose, R. F., and Poff, N. L. (1997). Ecological theory and community restoration ecology. Restor. Ecol. 5, 291–300. doi: 10.1046/j.1526-100X.1997.00543.x
Perring, M. P., Standish, R. J., Price, J. N., Craig, M. D., Erickson, T. E., Ruthrof, K. X., et al. (2015). Advances in restoration ecology: rising to the challenges of the coming decades. Ecosphere 6:131. doi: 10.1890/ES15-00121.1
Pilliod, D. S., and Welty, J. L. (2013). Land Treatment Digital Library. U.S. Geological Survey Data Series 806. doi: 10.3133/ds806
Pilliod, D. S., Welty, J. L., and Arkle, R. S. (2017a). Refining the cheatgrass–fire cycle in the Great Basin: precipitation timing and fine fuel composition predict wildfire trends. Ecol. Evol. 7, 8126–8145. doi: 10.1002/ece3.3414
Pilliod, D. S., Welty, J. L., and Toevs, G. R. (2017b). Seventy-five years of vegetation treatments on public rangelands in the Great Basin of North America. Rangelands 39, 1–9. doi: 10.1016/j.rala.2016.12.001
Prochazka, B. G., Coates, P. S., Ricca, M. A., Casazza, M. L., and Hull, J. M. (2017). Encounters with pinyon-juniper influence riskier movements in greater sage-grouse across the Great Basin. Rangeland Ecol. Manage. 70, 39–49. doi: 10.1016/j.rama.2016.07.004
Pyke, D. A. (2011). “Restoring and rehabilitating sagebrush habitats,” in Greater Sage-Grouse: Ecology and Conservation of a Landscape Species and Its Habitats, eds S. T. Knick and J. W. Connelly (Berkeley, CA: University of California Press), 531–548. doi: 10.1525/california/9780520267114.003.0024
Pyke, D. A., Chambers, J. C., Pellant, M., Knick, S. T., Miller, R. F., Beck, J. L., et al. (2015). Restoration Handbook for Sagebrush Steppe Ecosystems With Emphasis on Greater Sage-Grouse Habitat—Part 1. Concepts for Understanding and Applying Restoration. Report 1416, Reston, VA, 56. doi: 10.3133/cir1416
Reinhardt, J. R., Naugle, D. E., Maestas, J. D., Allred, B., Evans, J., and Falkowski, M. (2017). Next-generation restoration for sage-grouse: a framework for visualizing local conifer cutswithin a landscape context. Ecosphere 8:e01888. doi: 10.1002/ecs2.1888
Requena-Mullor, J. M., Maguire, K. C., Shinneman, D. J., and Caughlin, T. (2019). Integrating anthropogenic factors into regional-scale species distribution models — A novel application in the imperiled sagebrush biome. Glob. Change Biol. 25, 3844–3858. doi: 10.1111/gcb.14728
Resilience Alliance (2010). Assessing Resilience in Social-Ecological Systems: Workbook for Practitioners. Version 2.0. Available online at: http://www.resalliance.org/3871.php
Ricca, M. A., Coates, P. S., Gustafson, K. B., Brussee, B. E., Chambers, J. C., Espinosa, S. P., et al. (2018). A conservation planning tool for Greater Sage-grouse using indices of species distribution, resilience, and resistance. Ecol. Appl. 28, 878–896. doi: 10.1002/eap.1690
Rigge, M., Homer, C., Wylie, B., Gu, Y., Shi, H., Xian, G., et al. (2019). Using remote sensing to quantify ecosystem site potential community structure and deviation in the Great Basin, United States. Ecol. Indic. 96, 516–531. doi: 10.1016/j.ecolind.2018.09.037
Romme, W. H., Allen, C. D., Balley, J. D., Baker, W. L., Bestelmeyer, B. T., Brown, P. M., et al. (2009). Historical and modern disturbance regimes, stand structures, and landscape dynamics in pinon-juniper vegetation of the Western United States. Rangeland Ecol. Manage. 62, 203–222. doi: 10.2111/08-188R1.1
Roundy, B. A., Miller, R. F., Tausch, R. J., Young, K., Hulet, A., Rau, B., et al. (2014). Understory cover responses to piñon–juniper treatments across tree dominance gradients in the Great Basin. Rangeland Ecol. Manage. 67, 482–494. doi: 10.2111/REM-D-13-00018.1
Row, J. R., Doherty, K. E., Cross, T. B., Schwartz, M. K., Oyler-McCance, S. J., Naugle, D. E., et al. (2018). Quantifying functional connectivity: the role of breeding habitat, abundance, and landscape features on range-wide gene flow in sage-grouse. Evol. Appl. 11, 1305–1321. doi: 10.1111/eva.12627
Rowland, M. M., Wisdom, M. J., Suring, L. H., and Meinke, C. W. (2006). Greater sage-grouse as an umbrella species for sagebrush-associated vertebrates. Biol. Conserv. 129, 323–335. doi: 10.1016/j.biocon.2005.10.048
Runge, C. A., Withey, J. C., Naugle, D. E., Fargione, J. E., Helmstedt, K. J., Larsen, A. E., et al. (2019). Single species conservation as an umbrella for management of landscape threats. PLoS ONE 14:e0209619. doi: 10.1371/journal.pone.0209619
Sandford, C. P., Kohl, M. T., Messmer, T. A., Dahlgren, D. K., Cook, A., and Wing, B. R. (2017). Greater sage-grouse resource selection drives reproductive fitness under a conifer removal strategy. Rangeland Ecol. Manage. 70, 59–67. doi: 10.1016/j.rama.2016.09.002
Scheffer, M., Carpenter, S., Foley, J. A., Folke, C., and Walker, B. (2001). Catastrophic shifts in ecosystems. Nature 413, 591–596. doi: 10.1038/35098000
Schlaepfer, D. R., Lauenroth, W. K., and Bradford, J. B. (2014). Natural regeneration processes in big sagebrush (Artemisia tridentata). Rangeland Ecol. Manage. 67, 344–357. doi: 10.2111/REM-D-13-00079.1
Schroeder, M. A., Aldridge, C. L., Apa, A. D., Bohne, J. R., Braun, C. E., Bunnell, S. D., et al. (2004). Distribution of sage-grouse in North America. Condor 106, 363–376. doi: 10.1093/condor/106.2.363
Seastedt, T. R., Hobbs, R. J., and Suding, K. N. (2008). Management of novel ecosystems: are novel approaches required? Front. Ecol. Environ. 6, 547–553. doi: 10.1890/070046
Severson, J. P., Hagen, C. A., Maestas, J. D., Naugle, D. E., Forbes, J. T., and Reese, K. P. (2017a). Restoring Sage-grouse nesting habitat through removal of early successional conifer. Restor. Ecol. 25, 1026–1034. doi: 10.1111/rec.12524
Severson, J. P., Hagen, C. A., Tack, J. D., Maestas, J. D., Naugle, D. E., Forbes, J. T., et al. (2017b). Better living through conifer removal: a demographic analysis of sage-grouse vital rates. PLoS ONE 12:e0174347. doi: 10.1371/journal.pone.0174347
Shi, H., Rigge, M., Homer, C. G., Xian, G., Meyer, D. K., and Bunde, B. (2017). Historical cover trends in a sagebrush steppe ecosystem from 1985 to 2013: links with climate, disturbance, and management. Ecosystems 21, 913–929. doi: 10.1007/s10021-017-0191-3
Short, K. C., Finney, M. A., Scott, J. H., Gilbertson-Day, J. W., and Grenfell, I. C. (2016). Spatial Dataset of Probabilistic Wildfire Risk Components for the Conterminous United States. Fort Collins, CO: Forest Service Research Data Archive.
Shriver, R. K., Andrews, C. M., Arkle, R. S., Barnard, D. M., Duniway, M. C., Germino, M. J., et al. (2019). Transient population dynamics impede restoration and may promote ecosystem transformation after disturbance. Ecol. Lett. 22, 1357–1366. doi: 10.1111/ele.13291
Standish, R. J., Hobbs, R. J., Mayfield, M. M., Bestelmeyer, B. T., Suding, K. N., Battaglia, L. L., et al. (2014). Resilience in ecology: abstraction, distraction, or where the action is? Biol. Conserv. 177, 43–51. doi: 10.1016/j.biocon.2014.06.008
Stiver, S. J., Rinkes, E. T., Naugle, D. E., Makela, P. D., Nance, D. A., and Karl, J. W. (2015). Sage-Grouse Habitat Assessment Framework: A Multiscale Assessment Tool. Technical Reference 6710-1. Bureau of Land Management and Western Association of Fish and Wildlife Agencies, Denver, CO. doi: 10.5962/bhl.title.111572
Strand, E. K., Bunting, S. C., and Keefe, R. F. (2013). Influence of wildland fire along a successional gradient in sagebrush steppe and western juniper woodlands. Rangeland Ecol. Manage. 66, 667–679. doi: 10.2111/REM-D-13-00051.1
Stringham, T. K., Novak-Echenique, P., Snyder, D. K., Peterson, S., and Snyder, K. A. (2016). Disturbance response grouping of ecological sites increases utility of ecological sites and state-and-transition models for landscape scale planning in the Great Basin. Rangelands 38, 371–378. doi: 10.1016/j.rala.2016.10.006
Stringham, T. K., Snyder, K. A., Snyder, D. K., Lossing, S. S., Carr, C. A., and Stringham, B. J. (2018). Rainfall interception by singleleaf piñon and Utah juniper: implications for stand-level effective precipitation. Rangeland Ecol. Manage. 71, 327–335. doi: 10.1016/j.rama.2017.12.009
Sudduth, E. B., Hassett, B. A., Cada, P., and Bernhardt, E. S. (2011). Testing the field of dreams hypothesis: functional responses to urbanization and restoration in stream ecosystems. Ecol. Appl. 21, 1972–1988. doi: 10.1890/10-0653.1
Suding, K. N. (2011). Toward an era of restoration in ecology: successes, failures, and opportunities ahead. Annu. Rev. Ecol. Evol. Syst. 42, 465–487. doi: 10.1146/annurev-ecolsys-102710-145115
Suding, K. N., Gross, K. L., and Houseman, G. R. (2004). Alternative states and positive feedbacks in restoration ecology. Trends Ecol. Evol. 19, 46–53. doi: 10.1016/j.tree.2003.10.005
Suring, L. H., Wisdom, M. J., Tausch, R. J., Miller, R. F., Rowland, M. M., Schueck, L., et al. (2005). “Modeling threats to sagebrush and other shrubland communities,” in Habitat Threats in the Sagebrush Ecosystem; Methods of Regional Assessment and Applications in the Great Basin, eds M. J. Wisdom, M. M. Rowland, and L. H. Suring (Lawrence, KS: Alliance Communications Group), 114–149.
Taylor, R. L., Walker, B. L., Naugle, D. E., and Mills, L. S. (2012). Managing multiple vital rates to maximize greater sage-grouse population growth. J. Wildlife Manage. 76, 336–347. doi: 10.1002/jwmg.267
U S. Fish and Wildlife Service (2015b). Endangered and Threatened Wildlife and Plants; Withdrawal of Proposed Rule to List the Bi-State Distinct Population Segment of Greater Sage-Grouse and Designate Critical Habitat. U.S. Fish and Wildlife Service, Department of Interior. 80 FR 22827.
U.S. Fish Wildlife Service (2013). U.S. Fish and Wildlife Service. Greater Sage-grouse (Centrocercus urophasianus) Conservation Objectives: Final Report. U.S. Fish and Wildlife Service, Denver, CO. Available online at: http://www.fws.gov/greatersagegrouse/documents/COT-Report-with-Dear-Interested-Reader-Letter.pdf.
U.S. Fish and Wildlife Service (2015a). Endangered and Threatened Wildlife and Plants; 12-Month Finding on a Petition to List Greater Sage-Grouse (Centrocercus urophasianus) as an Endangered or Threatened Species. U.S. Fish and Wildlife Service, Department of Interior. 80 FR 59857.
WAFWA (2015). Greater Sage-Grouse Population Trends: An Analysis of Lek Count Databases 1965–2015. Western Association of Fish and Wildlife Agencies, Cheyenne, WY. Available online at: http://www.wafwa.org/Documents%20and%20Settings/37/Site%20Documents/News/Lek%20Trend%20Analysis%20final%208-14-15.pdf.
Keywords: Artemesia conservation planning, Centrocercus urophasianus, conifer expansion, sage-grouse, suitability, wildfire, umbrella species
Citation: Ricca MA and Coates PS (2020) Integrating Ecosystem Resilience and Resistance Into Decision Support Tools for Multi-Scale Population Management of a Sagebrush Indicator Species. Front. Ecol. Evol. 7:493. doi: 10.3389/fevo.2019.00493
Received: 04 July 2019; Accepted: 03 December 2019;
Published: 14 January 2020.
Edited by:
Samuel A. Cushman, United States Forest Service (USDA), United StatesReviewed by:
Mohammad Imam Hasan Reza, Independent Researcher, Chittagong, BangladeshJonah Henri Ratsimbazafy, Madagascar Primate Study and Research Group, Madagascar
Copyright © 2020 Ricca and Coates. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peter S. Coates, pcoates@usgs.gov