Plant Attributes that Drive Dispersal and Establishment Limitation in Tropical Agricultural Landscapes
by
 , ,
, ,
Marinés De la Peña-Domene
1,2,* ,
,
Cristina Martínez-Garza
1,
Luz María Ayestarán-Hernández
1 and
Henry F. Howe
3 1
Centro de Investigación en Biodiversidad y Conservación, Universidad Autónoma del Estado de Morelos, Cuernavaca, Morelos 62209, Mexico
2
Coordinación General de Adaptación al Cambio Climático del Instituto Nacional de Ecología y Cambio Climático, Ciudad de México 14210, Mexico
3
Department of Biological Sciences, University of Illinois at Chicago (m/c 066), Chicago, IL 60607, USA
*
Author to whom correspondence should be addressed.
Forests 2018, 9(10), 620; https://doi.org/10.3390/f9100620
Submission received: 3 August 2018
/
Revised: 30 September 2018
/
Accepted: 1 October 2018
/
Published: 10 October 2018
(This article belongs to the Special Issue The Forest Renewal Issue: New Problems and Findings in the Establishment of Tree Regeneration)
Abstract
:Factors that influence tropical-forest regeneration have been of interest across the tropics. We tested the degree of dispersal and establishment limitation of pioneer and non-pioneer tree species with different dispersal modes and seed sizes, using data on both seed fall and seedling establishment in primary forest, secondary forest, and pasture excluded from livestock. The study took place in a lowland tropical rain forest in southeastern Mexico. To calculate dispersal and establishment limitation, we used a density-weighted index that considers: (1) whether a seed or seedling of a given species has arrived in the sample area; and (2) the fraction of seeds or seedlings contributed by a given species relative to the total number of seeds or seedlings arriving at a sampling station. Dispersal limitation of non-pioneer species and animal-dispersed species decreased with succession. The secondary forest had less dispersal limitation for wind-dispersed pioneers than pasture, resulting in a dense aggregation of species with seeds dispersed by wind. Overall, establishment limitation differed between animal-dispersed and wind-dispersed species in the primary forest, and was negatively correlated with seed size. The low capacity of most species to arrive, germinate, and establish as seedlings in pastures slows succession back to forest. To overcome barriers to natural succession in pastures, transplanting seedlings of non-pioneer species is suggested because most of them show high dispersal and establishment limitation.
1. Introduction
Two of the critical barriers to plant recruitment are failure of seeds to reach a site (dispersal limitation), and failure of seedling germination, survival and growth (establishment limitation). Dispersal limitation strongly limits regeneration of many plant species [1]. Seed arrival is influenced by dispersal agents; for example, some seeds depend on the mobility of animals such as primates, large rodents and large birds that select specific foraging areas while avoiding open areas [2,3,4,5]. In contrast, dispersal by wind is often inefficient inside the forest because wind currents are blocked and seeds collide with standing vegetation [6]. Very few large seeds of wind-dispersed species travel more than 10 m from the forest edge [7]. However, in open or early successional areas, small wind-dispersed seeds may travel hundreds of meters (e.g., up to 1.4 km in a deforested valley in Scotland [8]). Therefore, dispersal limitation may be higher in pastures for large-seeded species than in forested habitats given that most tree species in tropical forest are dispersed by animals [9]. Many small seeds that do arrive are wind-dispersed.
While the arrival of seeds in a suitable site is a necessary first step for a plant species to be present, seed arrival does not guarantee seed germination and seedling establishment [10,11]. A variety of abiotic and biotic factors filter seedling establishment in any habitat. Plant attributes vary among environmental gradients, thereby, seed dispersal plays a role in shaping local species assemblages [12]. For example, in open pastures, high temperature, low humidity and strong competition with grasses depress seedling establishment and survival [13,14]. On the other hand, accumulation of soil nutrients and higher humidity under tree canopies favor seedling recruitment [15]; however, restricted light under trees alters competition among heterospecifics, constraining juvenile growth and survival of some species more than others [13,16,17,18]. After seed arrival, the establishment of seedlings depends largely on whether the conditions where the seed is deposited favor germination and seedling survival.
Seed size affects both seed dispersal and seedling survival. Small-seeded species (seeds <4 mm wide), mainly pioneers, produce and disperse more seeds than large-seeded non-pioneer species [19]. They also have higher rates of seed predation, especially by ants and small rodents [13,20,21,22,23]. Large-seeded, non-pioneer species (seeds >4 mm wide) are less susceptible to pathogens [24,25], and they experience lower predation rates in areas without a canopy because their natural predators rarely visit those areas [16,21]. Additionally, seedlings from large seeds are less susceptible to droughts and high temperatures than seedlings from smaller seeds [18,22]. The vast majority of seeds >4 mm wide in the Neotropics depend on animal vectors for dispersal, thereby overcoming what would otherwise be extreme dispersal limitation [26]. For some species, the rate of seedling establishment is so high that there is little difference between dispersed seeds and established seedlings. For others, the disparity between seed arrival and actual recruitment may be immense [26]. While many small seeds of pioneer species that arrive in open areas fail to survive, few larger-seeded non-pioneers with higher recruitment success are dispersed from forest to open areas. Overall, strategy differentiation among species contributes to the maintenance of diversity in forests across the tropics [27].
Our general objective was to test the degree of dispersal and establishment limitation of pioneer and non-pioneer tree species with different dispersal modes and seed sizes, using data on both seed fall and seedling establishment. Given higher dispersal and establishment limitation in pastures compared to forested habitats, we tested the hypothesis that small-seeded early successional tree species dispersed by animals show the lowest dispersal and establishment limitation in early succession, but both dispersal and establishment limitation of small-seeded species increase as succession progresses. By quantifying the dispersal and establishment limitation in primary forest, secondary forest, and pasture excluded from livestock, we can guide management and restoration actions in permanent agricultural landscapes.
2. Materials and Methods
2.1. Study Site
This study was conducted at Los Tuxtlas, Veracruz, Mexico (18°05′18″–18°45′ N; 94°35′–95°30′ W). The Los Tuxtlas forest is 3300 km2 in area and ranges in elevation from 200 to 1700 m above sea level. This volcanic region is characterized by acidic soils (pH 4.9) with a large clay component (48.5% clay) [28]. Mean annual precipitation was 4825 mm per year from 1997 to 2006, the rainy season is from June to February, the dry season is from March to May, and the mean annual temperature is 25 °C [29]. The formerly forested land in which our plots were established had been under intensive cattle ranching for at least 30 years on ground planted with native (Axonopus compressus (Sw.) P. Beauv. and Paspalum conjugatum P.J. Bergius) and exotic grasses (Cynodon dactylon (L.) Pers., Brachiaria brizantha (Hocht. ex A. Rich) Stapf., Brachiaria decumbens Stapf., and Panicum spp.). Soil depth ranged from 5 to 72 cm over volcanic rock [30]. The nearby forest has a closed canopy ~35 m high and holds ~300 native tree species [31,32,33]; Nectandra ambigens (S.F. Blake) C.K. Allen (Lauraceae) is the most common species in the canopy, while Pseudolmedia oxyphyllaria Donn. Sm. (Moraceae) and Astrocaryum mexicanum Liebm. ex Mart. (Arecaceae) are abundant in the mid-canopy and understory, respectively [34]. More than 150 fruit-eating birds and mammals inhabit these forests [35]. Secondary forests faced wild fires 10 years before the data were collected. These forests are now a closed canopy of a combination of early-, mid- and late-successional tree species.
2.2. Experimental Design
This study took place in the context of an experimental restoration project to recover dispersal processes in a permanent agricultural landscape. The experiment consisted of a 3 × 8 grid of 24 fenced plots (30 m × 30 m, each plot separated by 35 m) established in pasture of the agricultural colony of Ruiz Cortínez in July and August 2006. Additionally, one 90 m × 30 m plot was located in the secondary forest and another of the same dimensions was established in the primary forest. Further details on the management history of the study site and project details can be found in de la Peña-Domene et al. [36,37,38].
Seed rain was collected in four seed traps from each of the 24 pasture plots and in 12 seed traps in the primary forest and 12 in the secondary forest. The coordinates of each seed trap at the forested habitats were determined using a random number generator within the 90 m × 30 m plots. In each pasture plot, four seed traps were located at random, one in each quarter plot, for a total of 96 seed traps over a total area of 14,414 m2 of recently-fenced pasture. Each seed trap consisted of four 1-m poles of polyvinyl chloride (PVC) supporting a 1 m2 frame holding a cloth, with a rock in the middle to trap seeds. From January 2007 to February 2008, seed samples were collected monthly in pasture and twice a month in forested habitats to avoid germination in the higher-moisture forest sites. We cleaned, counted and identified the seeds to the species level at the State University of Morelos (UAEM), Mexico. Seed identification was done using the herbarium at Los Tuxtlas Biological Station and reference collections from previous studies [39]. A sample of seeds of each species was preserved and photographed as a reference collection at UAEM. To assess seedling recruitment (woody seedlings >10 cm high), we counted and identified all the seedlings found every four months from June 2007 to June 2008 in pasture, primary forest and secondary forest. Recruit samples were from the 5 m2 adjacent to seed trap in forested habitats. Each location with a seed trap and the adjacent area used to assess seedlings was named a “station” (i.e., 12 stations in primary forest, and 12 stations in secondary forest). In pasture, due to very low or null recruitment, a larger sample of 144 m2 was used. Hereafter, we limit our analysis to species for which we found both seeds and seedlings (see Table 1 for a complete overview of the seed). Bibliographical information was obtained to classify species according to life history (more light-demanding “pioneer” species versus more shade-tolerant “non-pioneer” species, and primary seed-dispersal agent (wind or animal), and average seed mass (Table 2) [31,32,33,40,41,42,43].
2.3. Data Analysis
To calculate dispersal and establishment limitation, we used a density-weighted index (see equations below) that considers: (1) whether a seed or seedling of a given species has arrived to the sample area (sensu Muller-Landau et al. [44]); and (2) the fraction of seeds or seedlings contributed by a given species relative to the total number of seeds or seedlings arriving at a sampling station (details on the index can be found in de la Peña-Domene et al. [38], of this Special Issue). This index can distinguish between scenarios when all the seeds (or seedlings) of a species occur at the same station from the scenario where they are more evenly distributed between stations.
The density-weighted dispersal limitation index is:
where a is the number of seed traps which received a seed of the given species, n is the total number of seed traps, si is the number of seeds of species i, and S is the total number of seeds across all species and traps.
Similarly, for establishment limitation, the density-weighted index is:
where r is the number of stations with both seeds and seedlings of species i, am is the number of seed traps receiving seeds of species i times the area in square meters, pi is the number of recruits of species i at the station, and P is the total number of recruits across all species.
To test the influence of each habitat, dispersal mode and life history on dispersal and establishment limitation, we ran generalized linear models with a negative binomial error distribution and a log link. The best fit models were Generalized Linear Models (GLMs) including habitat (primary forest, secondary forest and pasture), life history and the interaction of these two factors, and GLMs including habitat, dispersal mode and their interaction (when statistically significant). An example of the R code used to include interactions (1) and without the interactions (2) is presented below. To better understand the interactions in the models, we then ran a separate model for each habitat to test the differences between life history and dispersal agent groups within each habitat. Finally, we used linear regressions of log seed weight and dispersal and establishment limitation. Statistical tests were conducted in R statistical software (v. 3.4.3) (R Core Team [45]) and Statistica 7 (Statsoft [46]).
Example R code used to generate GLMs:
- mdlog1 <-glm.nb (DispInvWhole ~ habitat * Hist, data = DE)
- mdlog2 <-glm.nb (DispInvWhole ~ habitat + Hist, data = DE)
3. Results
3.1. General Results
We calculated dispersal and establishment limitation indices for 33 species, of which 12 species arrived to the primary forest, 15 species to the secondary forest and 13 species to pasture. Seed mass of these species varied from 0.0001 to 3.42 g. Only two species were recorded in all three habitats: the non-pioneer animal-dispersed tree Cupania glabra Sw. (Sapindaceae) and the pioneer animal-dispersed shrub Piper amalago L. (Piperaceae). Another five species were present in two of the habitats, and the remaining 26 species were present in only one habitat (Table 2). Density-weighted indices showed that of the total species evaluated, 60% (24 species) had high limitation of both dispersal and establishment (more than 66% of limitation). Only two species had medium limitation of both dispersal and establishment (from 33% to 66%). The remaining 14 species (35%) were more limited in dispersal than establishment or vice versa. On average, 78% of all species in the three habitats had high dispersal limitation (more than 67% limitation).
3.2. Dispersal Limitation
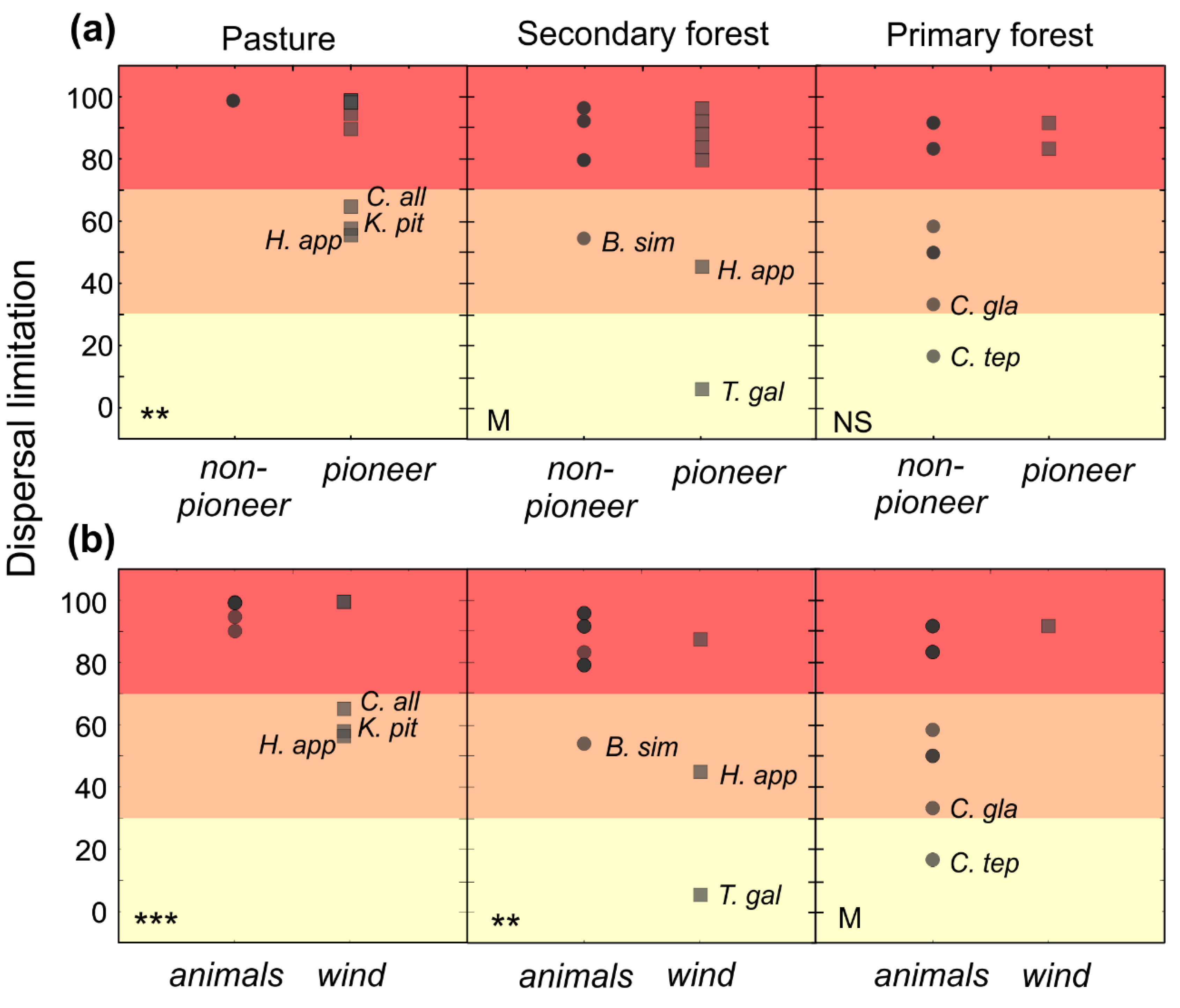
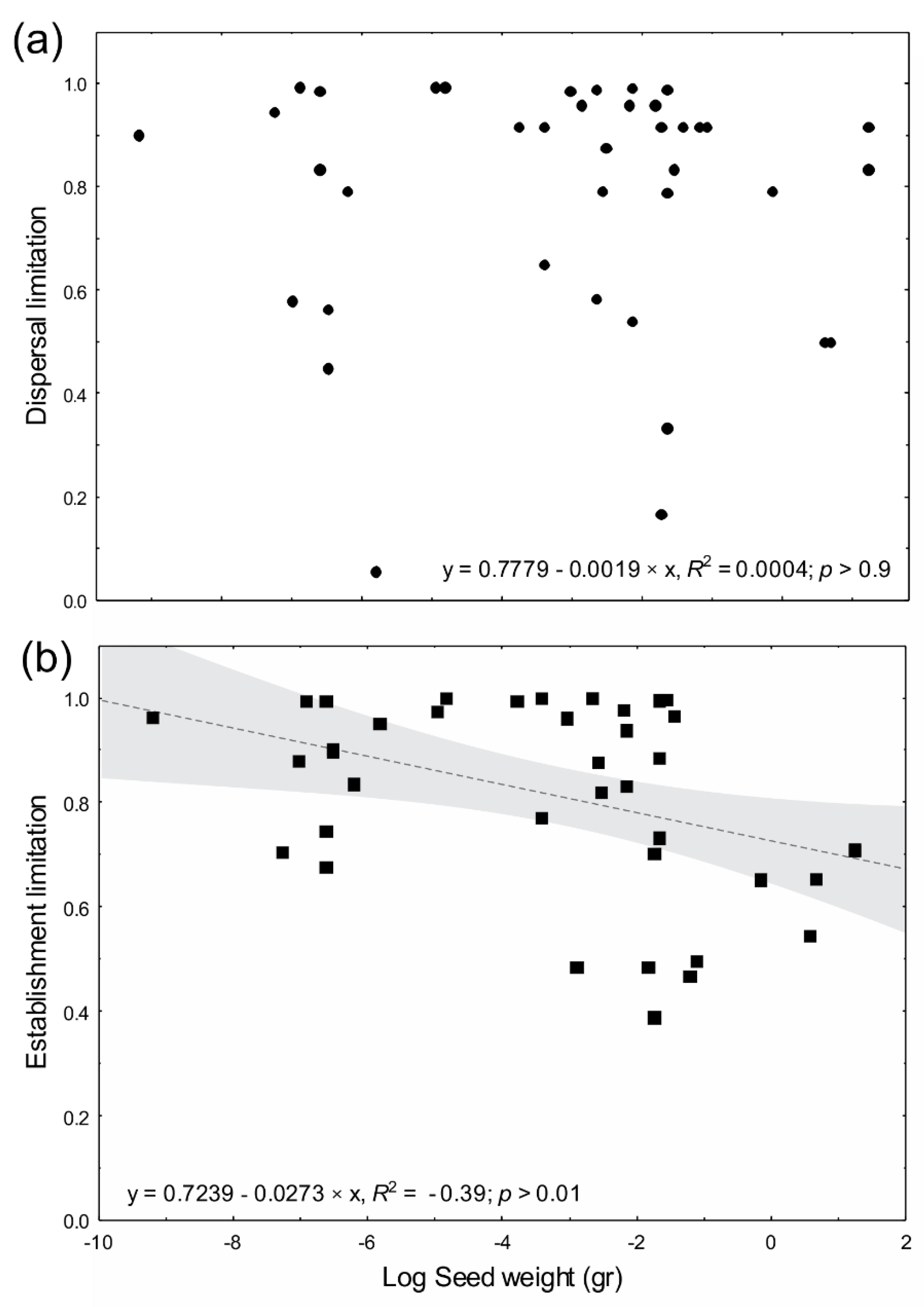
On average, over all habitats, dispersal limitation was significantly higher for pioneer species (80% ± 24%) than for non-pioneer species (77 ± 24; F = 2.61, p < 0.001) (Figure 1a and Table 3). Dispersal limitation was on average significantly higher for animal- (82% ± 35%) than wind-dispersed species (68% ± 31%; F = 2.21, p < 0.001) (Figure 1b and Table 3). When considering all habitats and species together (n = 33), dispersal limitation was not explained by seed size (R2 = 0.0004; p > 0.9, Figure 2a).
In pasture, non-pioneer species had significantly higher limitation than pioneer species (Figure 1a) and animal-dispersed species were significantly more limited than wind-dispersed species (Figure 1b and Table 3). Three wind-dispersed pioneer species showed intermediate dispersal limitation (60% ± 5%). The other 10 species in the pasture had extremely high dispersal limitation (>98% ± 3%).
In secondary forest, there was a marginal difference in dispersal limitation between life histories (Figure 1a and Table 3), and wind-dispersed species showed significantly higher dispersal limitation than animal-dispersed species (Figure 1c and Table 3). Only the wind-dispersed pioneer tree Trichospermum galeottii (Turcz.) Kosterm. (Malvaceae; 0.003 g.) was not dispersal limited (5% limitation). The wind-dispersed pioneer Heliocarpus appendiculatus Turcz. and animal-dispersed non-pioneer Bursera simaruba (L.) Sarg. had intermediate dispersal limitation (49% ± 6%), whereas the remaining 12 species were highly dispersal limited (87% ± 7%).
Dispersal limitation in the primary forest did not differ between life histories (Figure 1a and Table 3) and showed marginal differences between dispersal modes (Figure 1b and Table 3). Only the animal-dispersed palm Chamaedorea tepejilote Liebm. (Arecaceae; 0.175 g.) had low dispersal limitation (16%). Four other animal-dispersed non-pioneer tree species had intermediate dispersal limitation (48% ± 10%) and another three species showed high dispersal limitation (88% ± 4%) (Table 2).
3.3. Establishment Limitation
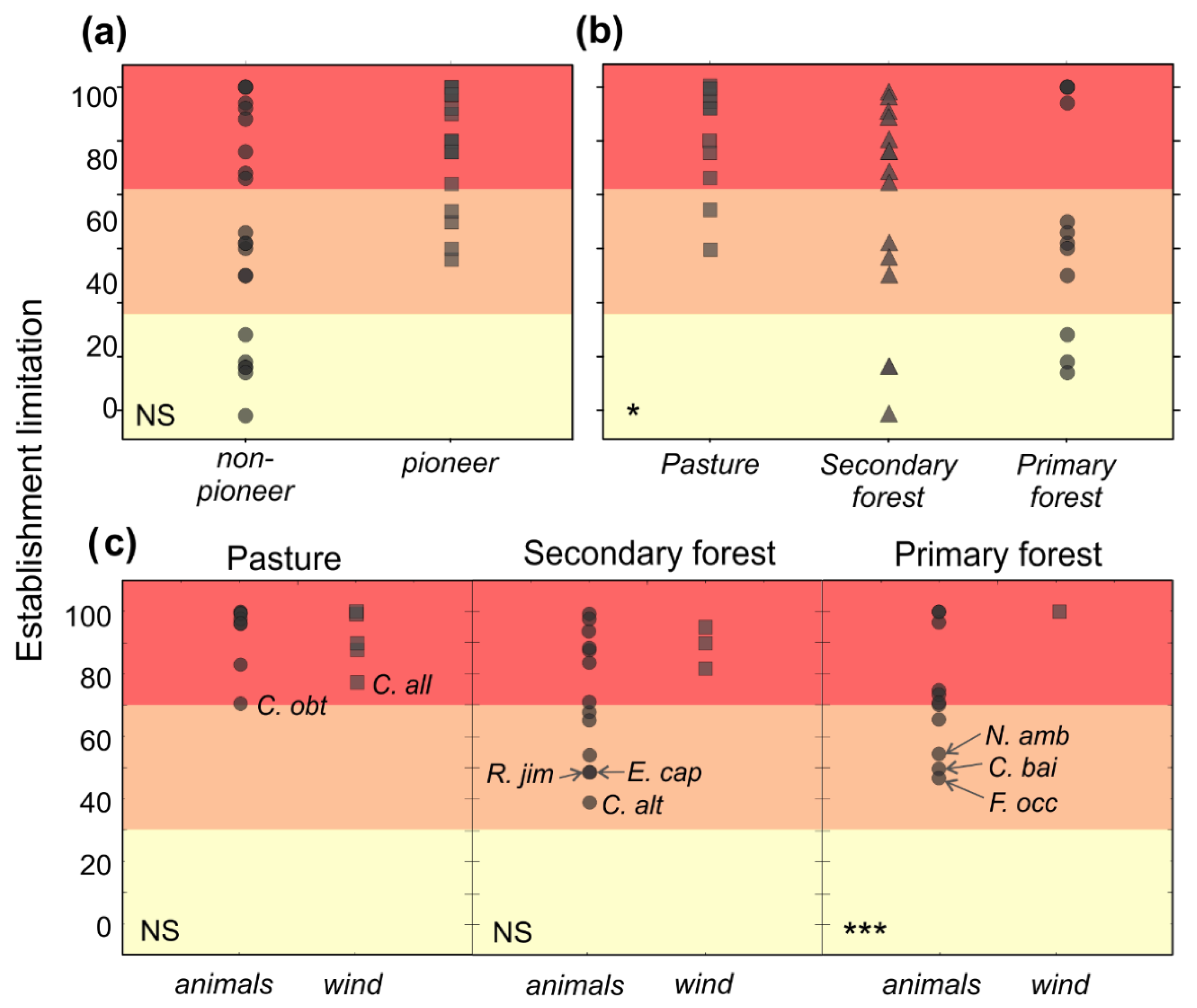
Establishment limitation was higher overall for pioneer (90% ± 11%) than for non-pioneer species (73% ± 2%) (Figure 3a and Table 3). Establishment limitation was also higher for wind-dispersed (91% ± 8%) than for animal-dispersed species (78% ± 19%) and statistically higher in the primary forest (Figure 3b and Table 3). In the primary forest, animal-dispersed species had significantly lower establishment limitation than wind-dispersed species (Figure 3b and Table 3). Four animal-dispersed non-pioneer species of the primary forest had intermediate establishment limitation (54% ± 8% limitation). Four species showed intermediate- to high-limitation (72% ± 2% limitation) and four were severely limited in establishment (99% ± 2% limitation).
Over all habitats and species (n = 33), seed size predicted establishment limitation (R2 = −0.39; p < 0.01) (Figure 2b). In pasture, establishment limitation was high for all species (92 ± 10% limitation). The pioneer species, Cecropia obtusifolia Bertol. (70%) and C. alliodora (Ruiz & Pav.) Oken (77%), showed the lowest establishment limitation. In the secondary forest, four animal-dispersed non-pioneer species had intermediate limitation (50% ± 11% limitation). Two more species showed intermediate- to high-establishment limitation (69% ± 2% limitation), and the remaining nine species were highly limited (91% ± 6% limitation) (Table 2).
4. Discussion
The distinction between dispersal limitation and establishment limitation is particularly important to predict the revegetation of areas formerly dedicated to agriculture. As successional age increased, we found a shift in dispersal limitation from non-pioneer species to pioneer species and from wind-dispersed species to animal-dispersed species. Establishment limitation was partially explained by seed weight, and was higher for wind-dispersed species and lower for animal-dispersed species as succession progressed.
4.1. Dispersal Limitation by Habitat
The degree of dispersal limitation is known to be variable in space, depending on local and landscape contexts [47]. Overall, our results showed that dispersal limitation was higher in pastures compared to forest habitats, as was expected. Other studies also show that dispersal limitation tends to be high for most species in primary forest [48], particularly, as we found, for wind-dispersed and pioneer species. It is widely known that wind-dispersed pioneer trees with small seeds often disperse better into open areas than animal-dispersed species [49,50,51]. We would expect this higher dispersal capacity of wind-dispersed pioneer species in open areas to result in their more overrepresentation in tropical pastures than in other habitats [52]. Our data do not support that assumption. In the open pasture, 38% of the species and 80% of the seeds were wind-dispersed, but in secondary forest, where there was an extremely high rate of seed fall (10,324 ± 5986 seeds m2/year), 20% of the species, yet 99% of the seeds, were wind-dispersed, 88% of which belonged to one wind-dispersed pioneer species, Trichospermum galeottii. This indicates that trees that produce high numbers of wind-dispersed seeds that are not in open areas might work as seed sinks of those species, leading to an over-representation of these seeds in the seed rain of secondary forests more than open pastures. Furthermore, secondary forests seem to have species with the colonization strategies of those in open areas but given the complex forest structure, the establishment of the arriving seeds is not favored.
During early succession in open pastures, seed availability is extremely low for most species. However, seed rain is known to have weak effects on the recovery of forest cover over a wide range of conditions, including when landscape-scale seed availability is either very high or very low [53]. In this study, only seeds of pioneer tree species dispersed by wind had dispersal limitation below 65%. Of those, only three pioneer species were even moderately successful in recruitment. Similarly, in pastures without reproductive trees near Las Cruces, Costa Rica, dispersal limitation shapes the initial composition, with small-seeded pioneer species driving abundance patterns [54]. Additionally, in sites where early and late pioneers establish, they may slow the influx of disperser-limited deep-forest trees for a century or more [55]. With extremely low seed input of most species except for very unsuccessful species for establishment, intensely managed cattle pastures have very little chances of recovering primary forest composition for a very long time, if ever.
4.2. Establishment Limitation by Habitat
A wide variety of mechanisms operating at multiple stages of a plant’s life contribute to the limitation of seedling establishment [56]. These include abiotic factors such as soil quality and moisture [30,57] as well as biotic factors including canopy cover [58] or seed and seedling predation by insects, pathogens or vertebrates [23,59]. Establishment limitation is important because dispersal limitation is extreme in highly diverse communities [60,61,62]. In the present study, the species with low dispersal limitation are not necessarily those with low establishment limitation. For example, dispersal limitation for wind-dispersed species was relatively low in open areas, yet establishment limitation for these species was remarkably high. In our study, establishment limitation differed between the two forest types and pasture, with the largest differences between secondary forest and pasture. The very high seed densities observed in secondary forest may increase seedling recruitment but may also induce negative density dependent mechanisms that act on high seedling densities, resulting in population regulation [10]. For these species, establishment might be more successful if dispersal is limited. The effect of recruitment limitation is that it allows inferior species to win sites in the absence of dominant competitors by forfeit [60]. During gap-phase regeneration in primary forests, community-wide recruitment limitation is very severe, even for pioneer species [63]. However, establishment limitation in our study was as high in pasture as in primary forest. Under recruitment limitation, many sites are won by “default” by species that are not the absolutely best competitor for the site.
Strong recruitment limitation does not prevent ultimate competitive exclusion in model communities, but it slows down the rate so much that the elimination of inferior competitors can be nearly infinitely delayed [63]. In pastures, low seedling survival slows natural succession. In this study, all species arriving in pastures showed high or extremely high establishment limitation. We found C. obtusifolia in 15% of the seed traps, ranging from 1 to 19 seeds in the first two years of pasture exclusions; a further study found 152,641 seeds in a period of six months in pastures after seven years of pasture exclusion [64]. Additionally, 40% of sites that had seeds of that species also had seedlings. After two years of abandonment of agricultural fields, in Marquéz de Comillas, Southeast Mexico, C. obtusifolia was the only species with seedlings, indicating strong establishment limitation for all other species [65]. Therefore, pastures that have been managed intensively for long periods of time face extreme dispersal and establishment limitations for most species that might not be overcome by unassisted regeneration alone.
4.3. Limitation by Life History, Dispersal Agent and Seed Size
Seed to seedling transitions are dependent on species traits, but also on successional stage of habitats [37]. Consistent with our hypothesis, the dispersal limitation of pioneer species was lower overall than non-pioneer species in early successional habitats, but higher in the primary forest. However, overall, pioneer species did not differ in establishment limitation compared to non-pioneer species. Contrary results were found in the seasonally moist tropical forest on Barro Colorado Island (BCI), Panama, a remnant primary forest island where the differences in establishment limitation paralleled differences in shade tolerance [44]; for pioneer species, factors affecting post-dispersal recruitment success were important contributors to seedling distribution patterns in forest gaps [66]. However, analyses of seedling recruitment in relation to seed abundance indicate that almost all tropical tree species, including pioneers, are likely to be strongly establishment-limited as well as seed-limited [44,62,67,68].
There is broad empirical evidence for the importance of dispersal mode in establishing the long-term community structure of tropical forests [69]. In primary tropical forest in peninsular Malaysia and in a 50-hectare tropical forest plot in Panama, the extent and scale of conspecific spatial aggregation is correlated with dispersal mode. This relationship holds for saplings as well as for mature trees [69]. Overall, we find no difference in dispersal limitation for species dispersed by wind or by animals. However, marginal differences are observed in establishment limitation where wind-dispersed species had higher limitation than animal-dispersed species. In fact, higher establishment limitation (but not dispersal limitation) was explained by lower seed weight, which may indicate that dispersal limitation is more related to animal behavior [70]. Nevertheless, even when wind-dispersed species have higher densities of dispersed seeds, they represent a small fraction of the diversity of tropical landscapes [9,71]. Four wind-dispersed pioneers were vastly over-represented in the pasture seed rain, but not as seedlings.
The relative strength of seed vs. establishment limitation might be expected to be largely dictated by seed size; we expected small-seeded pioneer species dispersed by animals to show the lowest dispersal limitation. Overall, we did not find this relationship between seed weight and dispersal limitation. In general, the high fecundity of small-seeded species should mean that they are less likely to be seed limited; however, small-seeded species have much lower seed-to-seedling transition probabilities than large-seeded species [36,67]. In restoration settings, in the landscape surrounding Las Cruces Biological Station, Costa Rica, seed rain shifted from a complete lack of tree seeds initially to a specific limitation on large-seeded, mature forest species over the first decade [54]. Accordingly, in deforested tropical mountains of Chulumani, Bolivia, seed limitation was particularly strong for large-seeded species [72]. Seed arrival of large-seeded species accounted for only 1% of the total seed pool in pastures of Los Tuxtlas, Mexico, in the same area as this study [7]. On the other hand, seed weight explained establishment limitation among all species. The consequence of seed size-mediated variation in seed and establishment limitation is an equilibrium competition colonization trade-off [73,74], which also acts to increase local species coexistence [44,75]. Overall, seeds cannot maximize both dispersibility and seedling survival, because they cannot be large and small at the same time.
4.4. Management Implications
The distinction between dispersal and establishment limitation can inform better management decisions. It is crucial to identify the specific mechanisms that limit each species before selecting restoration strategies [76]. When a species is dispersal-limited but has a high establishment probability (e.g., Cymbopetalum baillonii R.E. Fr. and Faramea occidentalis (L.) A. Rich. in the primary forest, Rollinia jimenezii Saff. and Chamaedorea alternans H. Wendl. in the secondary forest), management strategies for a given habitat can focus on facilitating the dispersal of the seeds, for example by seed sowing or direct seeding [19,77]. The same might be the case of animal-dispersed non-pioneer species in the primary forest, especially those that bear large seeds. Direct seeding may be less time-consuming, less logistically challenging and less costly than rearing seedlings [78]. Additionally, direct seeding avoids transplant shock, which can lead to higher stress and mortality of seedlings [79]. However, direct seeding for some species may have establishment issues that are much harsher than the planting of seedlings [80], and, therefore, selection of species for direct seeding and preparation of the site should be done carefully. Another approach would be to attract animal dispersers into a pasture by planting trees that offer cover and food resources in fruit crops [81]. In this case, the specific tree species arriving are not controlled, but the method becomes self-reinforcing as animal-dispersed species arrive and mature. If the main limitation is not seed availability, but rather establishment at a site (e.g., Trichospermum galeottii in the secondary forest and Bursera simaruba in the pasture), different management strategies are indicated. It is best to germinate seeds and grow seedlings to a viable size [82] in growing houses, and then plant them at the site [58]. This is likely the case for several species in the secondary forest, particularly wind-dispersed pioneers with small seeds. On the other hand, in pastures far away from forest (e.g., 50 m, see [7], and, in the case of this study, 100−500 m) where both dispersal and establishment limitations are high for most species, planting seedlings can overcome dispersal barriers [58]. Even if initial survival is challenging, over the years, plantings can improve microenvironmental conditions that help to increase seedling establishment [37]. If these plantings provide fleshy fruits, seed flow can also increase, most importantly, for large-seeded non-pioneer species [83]. Tree plantings provide a dual role in accelerating succession back to forest, but it is still unknown if animal dispersers are just attracted to plantings because they provide shelter, or if fruit-bearing trees are more attractive because they offer both food and shelter. Restoration plantings are always costly, but evaluating dispersal and establishment limitations can help allocate resources where they are more cost-effective.
5. Conclusions
Across all habitats, dispersal and establishment limitations were higher for pioneer species; dispersal limitation was higher for animal-dispersed species; and establishment limitation was higher for wind-dispersed species. In pastures, dispersal of non-pioneers was more limited, whereas, in the forest habitats, pioneers and non-pioneers showed similar dispersal limitation. Regarding dispersal modes, animal-dispersed species showed higher dispersal limitation in pasture, whereas wind-dispersed species were limited in secondary forest but similar dispersal limitation was found for animal and wind-dispersed species in primary forest. Establishment limitation was higher in the primary forest for all species. These results may inform management decisions to select restoration strategies for different species in the variety of habitats of agricultural landscapes.
Author Contributions
Conceptualization, M.P.-D., H.F.H. and C.M.-G.; Methodology, M.P.-D.; Validation, M.P.-D, L.M.A.-H. and C.M.-G.; Formal Analysis, M.P.-D, and C.M.-G.; Investigation, M.P.-D., H.F.H. and C.M.-G.; Resources, C.M.-G. and H.F.H.; Data Curation, M.P.-D., L.M.A.-H. and C.M.-G.; Writing—Original Draft Preparation, M.P.-D., C.M.-G.; Writing—Review and Editing, M.P.-D., C.M.-G. and H.F.H; Visualization, M.P.-D.; Supervision, C.M.-G.; Project Administration, C.M.-G. and H.F.H.; and Funding Acquisition, C.M.-G. and H.F.H.
Funding
The authors gratefully acknowledge financial and logistical support from the University of Illinois, Universidad Autónoma del Estado de Morelos, Universidad Nacional Autónoma de México, the Mexican Consejo Nacional de Ciencia y Tecnología (CONACyT) for a doctoral and postdoctoral fellowship, and the National Geographic Society and the National Science Foundation of the United States (DEB 0516259).
Acknowledgments
We are grateful to Rosamond Coates and Delfino Álvaro Campos Villanueva for institutional support; Eladio Velasco for expert field assistance; and the many students from ecology classes at Universidad Autónoma del Estado de Morelos, coordinated by Yuliana Urincho-Pantaleón, who helped process seed samples. We are grateful to Emiliano Cruz León, Miquel Gonzalez-Meler, Cesar Lozano Huerta, José Antonio Sierra Huelsz, Robin Foster, Emily Minor, Nigel Pitman, Rita Jiménez Rolland, Lynna Kiere and Corine Vriesendorp for discussions and constructive comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wijdeven, S.M.J.; Kuzee, M.E. Seed availability as a limiting factor in forest recovery processes in Costa Rica. Restor. Ecol. 2000, 8, 414–424. [Google Scholar] [CrossRef]
- Estrada, A.; Coates-Estrada, R. Diversity of neotropical migratory landbird species assemblages in forest fragments and man-made vegetation in Los Tuxtlas, Mexico. Biodivers. Conserv. 2005, 14, 1719–1734. [Google Scholar] [CrossRef]
- Estrada, A.; Coates-Estrada, R. Species composition and reproductive phenology of bats in a tropical landscape at Los Tuxtlas, Mexico. Camb. J. Online 2001, 17, 627–646. [Google Scholar] [CrossRef]
- Laborde, J.; Guevara, S.; Sanchez-Rios, G. Tree and shrub seed dispersal in pastures: The importance of rainforest trees outside forest fragments. EcoScience 2008, 15, 6–16. [Google Scholar] [CrossRef]
- Guevara, S.; Laborde, J. Monitoring seed dispersal at isolated standing trees in tropical pastures: Consequences for local species availability. Vegetatio 1993, 107/108, 319–338. [Google Scholar]
- Nathan, R.; Schurr, F.M.; Spiegel, O.; Steinitz, O.; Trakhtenbrot, A.; Tsoar, A. Mechanisms of long-distance seed dispersal. Trends Ecol. Evolut. 2008, 23, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Garza, C.; González-Montagut, R. Seed rain from forest fragments into tropical pastures in Los Tuxtlas, Mexico. Plant Ecol. 1999, 145, 255–265. [Google Scholar] [CrossRef]
- Bacles, C.F.E.; Lowe, A.J.; Ennos, R.A. Effective seed dispersal across a fragmented landscape. Science 2006, 311, 628. [Google Scholar] [CrossRef] [PubMed]
- Howe, H.F.; Smallwood, J. Ecology of seed dispersal. Annu. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Clark, C.J.; Poulsen, J.R.; Levey, D.J. Roles of seed and establishment limitation in determining patterns of afrotropical tree recruitment. PLoS ONE 2013, 8, e63330. [Google Scholar] [CrossRef] [PubMed]
- Reid, J.L.; Holl, K.D. Arrival ≠ survival. Restor. Ecol. 2013, 21, 153–155. [Google Scholar] [CrossRef]
- Weiher, E.; Freund, D.; Bunton, T.; Stefanski, A.; Lee, T.; Bentivenga, S. Advances, challenges and a developing synthesis of ecological community assembly theory. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 2403–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nepstad, D.C.; Uhl, C.; Serrao, E.A.S. Recuperation of a degraded Amazonian landscape: Forest recovery and agricultural restoration. Ambio 1991, 20, 248–255. [Google Scholar]
- Buschbacher, R.J. Tropical deforestation and pasture development. BioScience 1986, 36, 22–28. [Google Scholar] [CrossRef]
- Rhoades, C.C.; Eckert, G.E.; Coleman, D.C. Effect of pasture trees on soil nitrogen and organic matter: Implications for tropical montane forest restoration. Restor. Ecol. 1998, 6, 262–270. [Google Scholar] [CrossRef]
- Myster, R.W. Post-agricultural invasion, establishment, and growth of neotropical trees. Bot. Rev. 2004, 70, 381–402. [Google Scholar] [CrossRef]
- Turner, I.M. The Ecology of Trees in the Tropical Rain Forest; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Kitajima, K. Do shade-tolerant tropical tree seedlings depend longer on seed reserves? Functional growth analysis of three Bignoniaceae species. Funct. Ecol. 2002, 16, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Camargo, J.L.C.; Ferraz, I.D.K.; Imakawa, A.M. Rehabilitation of degraded areas of central Amazonia using direct sowing of forest tree seeds. Restor. Ecol. 2002, 10, 636–644. [Google Scholar] [CrossRef]
- García-Orth, X.; Martínez-Ramos, M. Seed dynamics of early and late successional tree species in tropical abandoned pastures: Seed burial as a way of evading predation. Restor. Ecol. 2008, 16, 435–443. [Google Scholar] [CrossRef]
- Jones, F.A.; Peterson, C.J.; Haines, B.L. Seed predation in neotropical pre-montane pastures: Site, distance, and species effects. Biotropica 2003, 35, 219–225. [Google Scholar] [CrossRef]
- Nepstad, D.; Uhl, C.; Serrão, A.E. Surmounting barriers to forest regeneration in abandoned highly degraded pastures: A case study from Paragominas, Pará, Brazil. In Alternatives to Deforestation: Steps toward Sustainable Use of the Amazon Rain Forest; Anderson, A.B., Ed.; Columbia University Press: New York, NY, USA, 1990; pp. 215–229. [Google Scholar]
- Myster, R.W. Vegetation dynamics of a permanent pasture plot in Puerto Rico. Biotropica 2003, 35, 422–428. [Google Scholar] [CrossRef]
- Sork, V.L. Effects of predation and light on seedling establishment in Gustavia superba. Ecology 1987, 68, 1341–1350. [Google Scholar] [CrossRef]
- Brokaw, N.V.L.; Scheiner, S.M. Species composition in gaps and structure of a tropical forest. Ecology 1989, 70, 538–541. [Google Scholar] [CrossRef]
- Terborgh, J.; Zhu, K.; Álvarez-Loayza, P.; Cornejo Valverde, F. How many seeds does it take to make a sapling? Ecology 2013, 95, 991–999. [Google Scholar] [CrossRef]
- Kraft, N.J.; Valencia, R.; Ackerly, D.D. Functional traits and niche-based tree community assembly in an Amazonian forest. Science 2008, 322, 580–582. [Google Scholar] [CrossRef] [PubMed]
- Guevara, S.; Laborde, D.; Sánchez-Ríos, G. Los Tuxtlas: El Paisaje de la Sierra; Instituto de Ecología, A.C.-Unión Europea: Xalapa, Mexico, 2004; p. 288. [Google Scholar]
- Soto, M.; Gama, L. La región de los tuxtlas: Climas. In Historia Natural de Los Tuxtlas; González Soriano, E., Dirzo, R., Vogt, C.R., Eds.; Universidad Nacional Autónoma de México, Instituto de Biología: Mexico City, Mexico, 1997; pp. 7–23. [Google Scholar]
- Tobon, W.; Martinez-Garza, C.; Campo, J. Soil responses to restoration of a tropical pasture in Veracruz, south-eastern Mexico. J. Trop. For. Sci. 2011, 23, 338–344. [Google Scholar]
- Ibarra-Manriquez, G.; Sinaca, S. Lista florística comentada de la Estación de Biología Tropical “Los Tuxtlas”, Veracruz, México. Rev. de Biol. Trop. 1995, 43, 75–115. [Google Scholar]
- Ibarra-Manriquez, G.; Sinaca, S. Lista florística comentada de la Estación de Biología Tropical “Los Tuxtlas”, Veracruz, México (Mimosaceae a Verbenaceae). Rev. de Biol. Trop. 1996, 44, 41–60. [Google Scholar]
- Ibarra-Manriquez, G.; Sinaca, S. Lista florística comentada de la Estación de Biología Tropical “Los Tuxtlas”, Veracruz, México (Violaceae a Zingiberaceae). Rev. de Biol. Trop. 1996, 44, 427–447. [Google Scholar]
- Bongers, F.; Popma, J.; Meave-del-Castillo, J.; Carabias, J. Structure and floristic composition on the lowland rain forest of Los Tuxtlas, Mexico. Vegetation 1988, 74, 55–80. [Google Scholar] [CrossRef]
- Estrada, A.; Coates-Estrada, R.; Meritt, D.; Montiel, S.; Curiel, D. Patterns of frugivore species richness and abundance in forest islands and in agricultural habitats at Los Tuxtlas, Mexico. Vegetatio 1993, 107/108, 245–257. [Google Scholar]
- De la Peña-Domene, M.; Howe, H.F.; Cruz-León, E.; Jiménez-Rolland, R.; Lozano-Huerta, C.; Martínez-Garza, C. Seed to seedling transitions in successional habitats across a tropical landscape. Oikos 2017, 126, 410–419. [Google Scholar] [CrossRef]
- de la Peña-Domene, M.; Martínez-Garza, C.; Howe, H.F. Early recruitment dynamics in tropical restoration. Ecol. Appl. 2013, 23, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- de la Peña-Domene, M. and Martínez-Garza, C. Integrating Density into Dispersal and Establishment Limitation Equations in Tropical Forests. Forests. 2018, 9, 570. [Google Scholar] [CrossRef]
- Martínez-Garza, C.; Flores-Palacios, A.; De La Peña-Domene, M.; Howe, H.F. Seed rain in a tropical agricultural landscape. J. Trop. Ecol. 2009, 25, 541–550. [Google Scholar] [CrossRef]
- Ibarra-Manriquez, G.; Ramos, M.M.; Oyama, K. Seedling functional types in a lowland rain forest in Mexico. Am. J. Bot. 2001, 88, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ramos, M. Claros, ciclos vitales de los árboles tropicales y la regeneración natural de las selvas altas perennifolias. In Investigaciones Sobre la Regeneración de las Selvas Altas en Veracruz, México; Amo, A.G.-P.S.d., Ed.; Editorial Alhambra Mexicana, S.A.: México D.F., Mexico, 1985; pp. 191–239. [Google Scholar]
- Popma, J.; Bongers, F.; Werger, M.J.A. Gap-dependance and leaf characteristics of trees in a tropical lowland rain-forest in Mexico. Oikos 1992, 63, 207–214. [Google Scholar] [CrossRef]
- Kew, R.B.G. Seed Information Database (sid), Version 7.1. Available online: http://data.kew.org/sid/ (accessed on 10 September 2011).
- Muller-Landau, H.C.; Wright, S.J.; Calderón, O.; Hubbell, S.P.; Foster, R.B. Assessing recruitment limitation: Concepts, methods and case-studies from a tropical forest. In Seed Dispersal and Frugivory; Levey, D.J., Silva, W.R., Galetti, M., Eds.; CAB International: Wallingford: Oxon, UK, 2002; pp. 35–53. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- “Statistica”, version 7; Data Analysis Software System; StatSoft Inc.: Tulsa, OK, USA, 2004.
- Caughlin, T.T.; Ferguson, J.M.; Lichstein, J.W.; Bunyavejchewin, S.; Levey, D.J. The importance of long-distance seed dispersal for the demography and distribution of a canopy tree species. Ecology 2013, 95, 952–962. [Google Scholar] [CrossRef]
- Norden, N.; Chave, J.; Belbenoit, P.; Caubere, A.; Chatelet, P.; Forget, P.M.; Riera, B.; Viers, J.; Thebaud, C. Interspecific variation in seedling responses to seed limitation and habitat conditions for 14 neotropical woody species. J. Ecol. 2009, 97, 186–197. [Google Scholar] [CrossRef]
- Cubina, A.; Aide, T.M. The effect of distance from forest edge on seed rain and soil seed bank in a tropical pasture. Biotropica 2001, 33, 260–267. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Pascarella, J.B.; Aide, T.M. Barriers to forest regeneration in an abandoned pasture in Puerto Rico. Restor. Ecol. 2000, 8, 350–360. [Google Scholar] [CrossRef]
- Vieira, D.L.M.; Scariot, A. Principles of natural regeneration of tropical dry forests for restoration. Restor. Ecol. 2006, 14, 11–20. [Google Scholar] [CrossRef]
- Posada, J.M.; Aide, T.M.; Cavelier, J. Cattle and weedy shrubs as restoration tools of tropical montane rainforest. Restor. Ecol. 2000, 8, 370–379. [Google Scholar] [CrossRef]
- Caughlin, T.T.; Elliott, S.; Lichstein, J.W. When does seed limitation matter for scaling up reforestation from patches to landscapes? Ecol. Appl. 2016, 26, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Reid, J.L.; Holl, K.D.; Zahawi, R.A. Seed dispersal limitations shift over time in tropical forest restoration. Ecol. Appl. 2015, 25, 1072–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finegan, B. Pattern and process in neotropical secondary rain forests: The first 100 years of succession. Trends Ecol. Evolut. 1996, 11, 119–124. [Google Scholar] [CrossRef]
- Schupp, E.; Milleron, T.; Russo, S. Dissemination limitation and the origin and maintenance of species-rich tropical forests. In Seed Dispersal and Frugivory: Ecology, Evolution and Conservation; Levey, D., Silva, W., Galetti, M., Eds.; CABI Publishing: Oxon, UK, 2002; pp. 19–33. [Google Scholar]
- Engelbrecht, B.M.J.; Wright, S.J.; De Steven, D. Survival and ecophysiology of tree seedlings during El Niño drought in a tropical moist forest in Panama. J. Trop. Ecol. 2002, 18, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Garza, C.; Bongers, F.; Poorter, L. Are functional traits good predictors of species performance in restoration plantings in tropical abandoned pastures? For. Ecol. Manag. 2013, 303, 35–45. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–527. [Google Scholar] [CrossRef]
- Hurtt, G.C.; Pacala, S.W. The consequences of recruitment limitation: reconciling chance, history and competitive differences between plants. J. Theor. Biol. 1995, 176, 1–12. [Google Scholar] [CrossRef]
- Ter Steege, H.; Pitman, N.C.A.; Sabatier, D.; Baraloto, C.; Salomão, R.P.; Guevara, J.E.; Phillips, O.L.; Castilho, C.V.; Magnusson, W.E.; Molino, J.-F.; et al. Hyperdominance in the Amazonian tree flora. Science 2013, 342, 1243092. [Google Scholar] [CrossRef] [PubMed]
- Terborgh, J.; Alvarez-Loayza, P.; Dexter, K.; Cornejo, F.; Carrasco, C. Decomposing dispersal limitation: Limits on fecundity or seed distribution? J. Ecol. 2011, 99, 935–944. [Google Scholar] [CrossRef]
- Hubbell, S.P.; Foster, R.B.; O’Brien, S.T.; Harms, K.E.; Condit, R.; Wechsler, B.; Wright, S.J.; de Lao, S.L. Light-gap disturbances, recruitment limitation, and tree diversity in a neotropical forest. Science 1999, 283, 554–557. [Google Scholar] [CrossRef] [PubMed]
- Popoca-Ortega, L.I. Lluvia de semillas en parcelas de restauración ecológica en la selva tropical de Los Tuxtlas, Veracruz, México. Bachelor Thesis, Universidad Autónoma del Estado de Morelos, Cuernavaca, Morelos, Mexico, 2016. [Google Scholar]
- Martínez-Ramos, M.; Pingarroni, A.; Rodríguez-Velázquez, J.; Toledo-Chelala, L.; Zermeño-Hernández, I.; Bongers, F. Natural forest regeneration and ecological restoration in human-modified tropical landscapes. Biotropica 2016, 48, 745–757. [Google Scholar] [CrossRef]
- Dalling, J.W.; Muller-Landau, H.C.; Wright, S.J.; Hubbell, S.P. Role of dispersal in the recruitment limitation of neotropical pioneer species. J. Ecol. 2002, 90, 714–727. [Google Scholar] [CrossRef] [Green Version]
- Harms, K.E.; Wright, S.J.; Calderon, O.; Hernandez, A.; Herre, E.A. Pervasive density-dependent recruitment enhances seedling diversity in a tropical forest. Nature 2000, 404, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Dalling, J.W.; Hubbell, S.P. Seed size, growth rate and gap microsite conditions as determinants of recruitment success for pioneer species. J. Ecol. 2002, 90, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Seidler, T.G.; Plotkin, J.B. Seed dispersal and spatial pattern in tropical trees. PLoS Biol. 2006, 4, e344. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, N.J.; Howe, H.F. Low recruitment of trees dispersed by animals in African forest fragments. Conserv. Biol. 2001, 15, 1733–1741. [Google Scholar] [CrossRef]
- Holl, K.D. Factors limiting tropical rain forest regeneration in abandoned pasture: Seed rain, seed germination, microclimate, and soil. Biotropica 1999, 31, 229–242. [Google Scholar] [CrossRef]
- Saavedra, F.; Hensen, I.; Schleuning, M. Deforested habitats lack seeds of late-successional and large-seeded plant species in tropical montane forests. Appl. Veg. Sci. 2015, 18, 603–612. [Google Scholar] [CrossRef]
- Hastings, A. Disturbance, coexistence, history, and competition for space. Theor. Popul. Biol. 1980, 18, 363–373. [Google Scholar] [CrossRef]
- Tilman, D. Competition and biodiversity in spatially structured habitats. Ecology 1994, 75, 2–16. [Google Scholar] [CrossRef]
- Chave, J.; Muller-Landau, H.C.; Levin, S.A. Comparing classical community models: Theoretical consequences for patterns of diversity. Am. Nat. 2002, 159, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Aradottir, A.L.; Halldorsson, G. Colonization of woodland species during restoration: Seed or safe site limitation? Restor. Ecol. 2018, 26, 738. [Google Scholar] [CrossRef]
- Turnbull, L.A.; Crawley, M.J.; Rees, M. Are plant populations seed-limited? A review of seed sowing experiments. Oikos 2000, 88, 225–238. [Google Scholar] [CrossRef]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of degraded tropical forest landscapes. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Luna, A.; Martínez-Garza, C. Performance of 15 tropical tree species recruited or transplanted on restoration settings. Bot. Sci. 2016, 94, 773. [Google Scholar] [CrossRef] [Green Version]
- Grossnickle, S.C.; Ivetić, V. Direct seeding in reforestation—A field performance review. Reforesta 2017, 4, 94–142. [Google Scholar] [CrossRef]
- de la Peña-Domene, M.; Martínez-Garza, C.; Palmas-Pérez, S.; Rivas-Alonso, E.; Howe, H.F. Roles of birds and bats in early tropical-forest restoration. PLoS ONE 2014, 9, e104656. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Garza, C.; Tobon, W.; Campo, J.; Howe, H.F. Drought mortality of tree seedlings in an eroded tropical pasture. Land Degrad. Dev. 2013, 24, 287–295. [Google Scholar] [CrossRef]
- de la Peña-Domene, M.; Minor, E.S.; Howe, H.F. Restored connectivity facilitates recruitment by an endemic large-seeded tree in a fragmented tropical landscape. Ecology 2016, 97, 2511–2517. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Plotted raw data of dispersal limitation by: (a) habitat and life history; and (b) habitat and dispersal mode. Each circle or square represents one of the 33 tropical tree species. The generalized linear models (GLM) showed significance for the interaction of: (a) habitat and life history; and (b) habitat and dispersal mode. Statistical significance between (a) pioneer and non-pioneer species and (b) animal- and wind-dispersed species within each habitat are indicated with asterisks (** p < 0.01, *** p < 0.001), M for marginal significance and NS for non-significant. Limitation from 0% to 33% was classified as low (yellow), from 34% to 66% as intermediate (orange) and from 67% to 100% as high (red).
Figure 1.
Plotted raw data of dispersal limitation by: (a) habitat and life history; and (b) habitat and dispersal mode. Each circle or square represents one of the 33 tropical tree species. The generalized linear models (GLM) showed significance for the interaction of: (a) habitat and life history; and (b) habitat and dispersal mode. Statistical significance between (a) pioneer and non-pioneer species and (b) animal- and wind-dispersed species within each habitat are indicated with asterisks (** p < 0.01, *** p < 0.001), M for marginal significance and NS for non-significant. Limitation from 0% to 33% was classified as low (yellow), from 34% to 66% as intermediate (orange) and from 67% to 100% as high (red).

Figure 2.
Relationship between: (a) seed weight and dispersal limitation; and (b) seed weight and establishment limitation (R2 = −0.39; p > 0.01) with regression line; gray shading shows overall relationships fitted with a linear model.
Figure 2.
Relationship between: (a) seed weight and dispersal limitation; and (b) seed weight and establishment limitation (R2 = −0.39; p > 0.01) with regression line; gray shading shows overall relationships fitted with a linear model.

Figure 3.
Plotted raw data of establishment limitation by: (a) life history; (b) habitat; and (c) habitat by dispersal mode among 33 tropical tree species. Different symbols (circles, squares or triangles) indicate different categories. The GLM model did not show significance for the interaction of habitat and life history, therefore the factors are shown independently marking the statistical significance between habitats and between life histories among all species. The interaction between habitat and dispersal mode was significant. Statistical significance between wind- and animal-dispersed species of each habitat is indicated with asterisks when significant (* p < 0.05, *** p < 0.001), M for marginal significance and NS for non-significant differences. Limitation from 0% to 33% was classified as low (yellow), from 34% to 66% as intermediate (orange) and from 67% to 100% as high (red).
Figure 3.
Plotted raw data of establishment limitation by: (a) life history; (b) habitat; and (c) habitat by dispersal mode among 33 tropical tree species. Different symbols (circles, squares or triangles) indicate different categories. The GLM model did not show significance for the interaction of habitat and life history, therefore the factors are shown independently marking the statistical significance between habitats and between life histories among all species. The interaction between habitat and dispersal mode was significant. Statistical significance between wind- and animal-dispersed species of each habitat is indicated with asterisks when significant (* p < 0.05, *** p < 0.001), M for marginal significance and NS for non-significant differences. Limitation from 0% to 33% was classified as low (yellow), from 34% to 66% as intermediate (orange) and from 67% to 100% as high (red).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Total number of species and individuals found as seeds and seedlings from 2007 to 2008 in each of three habitats in a tropical agricultural landscape of Los Tuxtlas, Mexico.
Table 1.
Total number of species and individuals found as seeds and seedlings from 2007 to 2008 in each of three habitats in a tropical agricultural landscape of Los Tuxtlas, Mexico.
| Habitat | Seeds | Seedlings | ||
|---|---|---|---|---|
| Species | Seeds | Species | Seedlings | |
| Pasture | 128 | 29,296 | 35 | 292 |
| Secondary forest | 115 | 81,725 | 35 | 261 |
| Primary forest | 162 | 6160 | 19 | 593 |
Table 2.
Families, life history, dispersal mode, seed mass (g) and, dispersal and establishment limitation, of species registered in the seed rain and recruited in primary and secondary forest and pasture in Los Tuxtlas, Veracruz categorized by habitat.
Table 2.
Families, life history, dispersal mode, seed mass (g) and, dispersal and establishment limitation, of species registered in the seed rain and recruited in primary and secondary forest and pasture in Los Tuxtlas, Veracruz categorized by habitat.
| Species | Family | Life History | Dispersal Mode | Seed Mass | Dispersal Limitation | Establishment Limitation |
|---|---|---|---|---|---|---|
| PRIMARY FOREST | ||||||
| Astrocaryum mexicanum Liebm. ex Mart. | ARECACEAE | Non-pioneer | Animal | 3.422 | 0.833 | 0.706 |
| Cymbopetalum baillonii R.E. Fr. | ANNONACEAE | Non-pioneer | Animal | 0.3335 | 0.917 | 0.495 |
| Cupania glabra Sw. | SAPINDACEAE | Non-pioneer | Animal | 0.19 | 0.333 | 0.731 |
| Chamaedorea tepejilote Liebm. | ARECACEAE | Non-pioneer | Animal | 0.175 | 0.167 | 0.701 |
| Faramea occidentalis (L.) A. Rich. | RUBIACEAE | Non-pioneer | Animal | 0.3 | 0.917 | 0.466 |
| Nectandra ambigens (S.F. Blake) C.K. Allen | LAURACEAE | Non-pioneer | Animal | 1.8 | 0.5 | 0.542 |
| Notopleura chapensis (Steyerm.) C.M. Taylor | RUBIACEAE | Non-pioneer | Animal | 0.21 | 0.833 | 0.996 |
| Trichilia martiana C. DC. | MELIACEAE | Non-pioneer | Animal | 0.2364 | 0.917 | 0.966 |
| Virola guatemalensis (Hemsl.) Warb. | MYRISTICACEAE | Non-pioneer | Animal | 0.1962 | 0.5 | 0.653 |
| Stemmadenia donnell-smithii (Rose) Woodson | APOCYNACEAE | Pioneer | Animal | 0.07 | 0.583 | 0.999 |
| Piper amalago L. | PIPERACEAE | Pioneer | Animal | 0.0015 | 0.833 | 0.746 |
| Cordia alliodora (Ruiz & Pav.) Oken | BORAGINACEAE | Pioneer | Wind | 0.033 | 0.917 | 0.999 |
| SECONDARY FOREST | ||||||
| Astrocaryum mexicanum Liebm. ex Mart. | ARECACEAE | Non-pioneer | Animal | 3.422 | 0.917 | 0.71 |
| Bursera simaruba (L.) Sarg. | BURSERACEAE | Non-pioneer | Animal | 0.117 | 0.54 | 0.938 |
| Cojoba arborea (L.) Britton & Rose | FABACEAE | Non-Pioneer | Animal | 0.85 | 0.791 | 0.652 |
| Cupania glabra Sw. | SAPINDACEAE | Non-pioneer | Animal | 0.19 | 0.789 | 0.884 |
| Chamaedorea alternans H. Wendl. | ARECACEAE | Non-pioneer | Animal | 0.175 | 0.917 | 0.387 |
| Eugenia capuli (Schltdl. & Cham.) Hook. & Arn. | MYRTACEAE | Non-pioneer | Animal | 0.056 | 0.958 | 0.484 |
| Psychotria veracruzensis Lorence & Dwyer | RUBIACEAE | Non-pioneer | Animal | 0.002 | 0.792 | 0.835 |
| Rollinia jimenezii Saff. | ANNONACEAE | Non-pioneer | Animal | 0.16 | 0.958 | 0.484 |
| Carica papaya L. | CARICACEAE | Pioneer | Animal | 0.023 | 0.917 | 0.992 |
| Hampea nutricia Fryxell | MALVACEAE | Pioneer | Animal | 0.11 | 0.958 | 0.976 |
| Piper amalago L. | PIPERACEAE | Pioneer | Animal | 0.0015 | 0.833 | 0.677 |
| Cnidoscolus multilobus (Pax) I.M. Johnst. | EUPHORBIACEAE | pioneer | Animal | 0.08 | 0.875 | 0.817 |
| Heliocarpus appendiculatus Turcz. | MALVACEAE | Pioneer | Wind | 0.0015 | 0.45 | 0.901 |
| Trichospermum galeottii (Turcz.) Kosterm. | MALVACEAE | Pioneer | Wind | 0.003 | 0.056 | 0.95 |
| Vachellia cornigera (L.) Seigler & Ebinger | FABACEAE | Pioneer | Wind | 0.076 | 0.792 | 0.876 |
| PASTURE | ||||||
| Bursera simaruba (L.) Sarg. | BURSERACEAE | Non-pioneer | Animal | 0.117 | 0.992 | 0.829 |
| Cupania glabra Sw. | SAPINDACEAE | Non-pioneer | Animal | 0.19 | 0.99 | 0.995 |
| Tetrorchidium rotundatum Standl. | EUPHORBIACEAE | Non-pioneer | Animal | 0.048 | 0.987 | 0.961 |
| Cecropia obtusifolia Bertol. | URTICACEAE | Pioneer | Animal | 0.001 | 0.946 | 0.704 |
| Conostegia xalapensis (Bonpl.) D. Don ex DC. | MELASTOMATACEAE | Pioneer | Animal | 0.0001 | 0.9 | 0.962 |
| Piper amalago L. | PIPERACEAE | Pioneer | Animal | 0.0015 | 0.988 | 0.992 |
| Stemmadenia donnell-smithii (Rose) Woodson | APOCYNACEAE | Pioneer | Animal | 0.07 | 0.989 | 0.998 |
| Trema micrantha (L.) Blume | CANNABACEAE | Pioneer | Animal | 0.007 | 0.994 | 0.975 |
| Cordia alliodora (Ruiz & Pav.) Oken | BORAGINACEAE | Pioneer | Wind | 0.033 | 0.651 | 0.771 |
| Koanophyllon pittieri (Klatt) R.M. King & H. Robinson | ASTERACEAE | Pioneer | Wind | 0.0009 | 0.579 | 0.877 |
| Heliocarpus appendiculatus Turcz. | MALVACEAE | Pioneer | Wind | 0.0015 | 0.563 | 0.897 |
| Heliocarpus donnellsmithii Rose | MALVACEAE | Pioneer | Wind | 0.001 | 0.994 | 0.993 |
| Ochroma pyramidale (Cav. ex Lam.) Urb. | MALVACEAE | Pioneer | Wind | 0.008 | 0.994 | 0.998 |
Table 3.
Summary of results of the generalized linear models for each response variable analyzed (dispersal and establishment limitation) in the interaction of habitat (Primary Forest (PrimFor), Secondary Forest (SecFor) by life history (Hist) and habitat by dispersal mode (Disp)).
Table 3.
Summary of results of the generalized linear models for each response variable analyzed (dispersal and establishment limitation) in the interaction of habitat (Primary Forest (PrimFor), Secondary Forest (SecFor) by life history (Hist) and habitat by dispersal mode (Disp)).
| Interaction | Dispersal Limitation | Establishment Limitation | ||||||
|---|---|---|---|---|---|---|---|---|
| Habitat by Life History | ||||||||
| Estimate | Error | z Value | p Value | Estimate | Error | z Value | p Value | |
| (Intercept) | 2.33 | 0.59 | 3.95 | <0.001 | 8.46 | 2.86 | 2.96 | 0.003 |
| Habitat-Primary forest | 3.52 | 0.67 | 5.28 | <0.001 | 8.10 | 4.19 | 1.93 | 0.05 |
| Habitat-Secondary forest | 2.78 | 0.68 | 4.06 | <0.001 | 9.65 | 4.70 | 2.05 | 0.04 |
| Life history-pioneer | 2.61 | 0.67 | 3.91 | <0.001 | 0.66 | 3.32 | 0.20 | 0.84 |
| Habitat-PrimFor: Hist-pioneer | −3.64 | 1.01 | −3.60 | <0.001 | −5.94 | 6.49 | −0.91 | 0.36 |
| Habitat-SecFor: Hist-pioneer | −2.01 | 0.84 | −2.40 | 0.02 | −8.01 | 5.54 | −1.45 | 0.14 |
| Habitat Type by Dispersal Mode | ||||||||
| Estimate | Error | z Value | p Value | Estimate | Error | z Value | p Value | |
| (Intercept) | 3.29 | 0.31 | 10.31 | <0.001 | 8.50 | 1.70 | 5.014 | <0.001 |
| Habitat-Primary forest | 2.52 | 0.41 | 6.08 | <0.001 | 8.00 | 3.27 | 2.433 | 0.01 |
| Habitat-Secondary forest | 1.74 | 0.40 | 4.27 | <0.001 | 7.54 | 3.12 | 2.420 | 0.01 |
| Dispersal mode-wind | 2.21 | 0.50 | 4.36 | <0.001 | 1.09 | 2.97 | 0.367 | 0.71 |
| Habitat-PrimFor: Disp-wind | −3.60 | 1.05 | −3.41 | <0.001 | −16.60 | 4.14 | −4.005 | <0.001 |
| Habitat-SecFor: Disp-wind | −0.95 | 0.76 | −1.25 | 0.2 | −6.65 | 5.22 | −1.274 | 0.20 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Peña-Domene, M.D.l.; Martínez-Garza, C.; Ayestarán-Hernández, L.M.; Howe, H.F. Plant Attributes that Drive Dispersal and Establishment Limitation in Tropical Agricultural Landscapes. Forests 2018, 9, 620. https://doi.org/10.3390/f9100620
AMA Style
Peña-Domene MDl, Martínez-Garza C, Ayestarán-Hernández LM, Howe HF. Plant Attributes that Drive Dispersal and Establishment Limitation in Tropical Agricultural Landscapes. Forests. 2018; 9(10):620. https://doi.org/10.3390/f9100620
Chicago/Turabian StylePeña-Domene, Marinés De la, Cristina Martínez-Garza, Luz María Ayestarán-Hernández, and Henry F. Howe. 2018. "Plant Attributes that Drive Dispersal and Establishment Limitation in Tropical Agricultural Landscapes" Forests 9, no. 10: 620. https://doi.org/10.3390/f9100620
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.