Effects of Graphene on Bacterial Community Diversity and Soil Environments of Haplic Cambisols in Northeast China


School of Forestry, Northeast Forestry University, 26 Hexing Road, Harbin 150040, China
*
Author to whom correspondence should be addressed.
Forests 2018, 9(11), 677; https://doi.org/10.3390/f9110677
Submission received: 27 September 2018
/
Revised: 25 October 2018
/
Accepted: 26 October 2018
/
Published: 28 October 2018
(This article belongs to the Section Forest Ecology and Management)
Abstract
:Graphene is the thinnest and toughest two-dimensional nanomaterial yet discovered. However, it inevitably enters the biosphere, where it may pose potential risks to ecosystems. We investigated the impact of applied graphene concentrations on bacterial community diversity, physicochemical properties, and enzyme activities of Haplic Cambisols, the zonal soil of Northeastern China. Soils receiving 0, 10, 100, or 1000 mg kg−1 of graphene were incubated for 7, 15, 30, 60, or 90 days. Adding graphene significantly increased the community richness and diversity index of the bacterial community in Haplic Cambisols, as well as their abundances, but this impact varied with graphene concentration and incubation time. Compared with 0 mg kg−1 of graphene applied, soil bacteria abundance and diversity increased significantly during early stages of incubation (i.e., 7 and 15 days) under different concentrations of graphene, and was inhibited or remained unchanged by a longer incubation time, reaching a minima at 60 days but then following an upward trend. Graphene treatments influenced the bacterial community structure and metabolic function in Haplic Cambisols, and the bacterial community’s metabolic regulation mechanism varied with both incubation time and graphene concentration. The rank order of bacterial similarity in soils treated with graphene was 15 > 7 > 30 > 60 > 90 days. Throughout the incubation periods, except for a few unidentified bacteria, the relative abundances of Proteobacteria and Acidobacteria in the soil samples were the highest, with the number of Pseudomonas of Proteobacteria being particularly large. The rank order of bacterial abundance at the phylum level in Haplic Cambisols was 15 > 7 > 30 > 90 > 60 days. Graphene also influenced bacterial community diversity by affecting several key soil environmental factors, such as organic matter and hydrolytic nitrogen contents, as well as urease and catalase activities.
1. Introduction
Graphene is the thinnest and toughest two-dimensional nanomaterial yet discovered, consisting of a dense layer of carbon atoms wrapped in a honeycomb crystal lattice that features the most stable benzene six-membered ring structural unit [1,2]. Graphene, the basic structural unit of fullerenes, carbon nanotubes, and graphite, has a thickness of only 0.34 nm, a carrier mobility of 15,000 cm2 V−1 s−1, a thermal conductivity of 5000 W m−1 K−1, and its specific surface area is 2630 m2 g−1 [3,4]. This special structure endows it with many excellent properties in optical, electrical, and thermal applications, as well as mechanical strength [1,5,6]. Not surprisingly, graphene applications have made tremendous progress, in environmental applications, biomedicine, materials, electronics, optics, energy, and other industries [7,8,9]. However, graphene and its derivatives—like other nanomaterials—are stable in the environment and thus not easily degraded. Their widespread application, on the one hand, improves the daily quality of human life but, on the other hand, graphene inevitably enters the biosphere, where it may pose potential risks to ecosystems. Therefore, it is vital that we understand graphene’s behavior in the environment, and whether it induces potential toxicity or risks to organisms.
Soil underpins modern agriculture and forestry [10]. Key soil biological properties, namely microbial community structure and enzyme activity, are important indicators for measuring the effects of pollutants or foreign substances on the biological activity and metabolism ability of contaminated soils [11,12,13], including the impact of graphene on soil environments [14]. Nonetheless, there is considerable debate over whether and how graphene affects soil microbial community and enzyme activity. Some studies reported that graphene could change the physiological metabolism of soil microorganisms to a certain extent. Under pure incubation conditions, graphene decreased the survival of bacteria by damaging their cell membranes [15] in addition to inhibiting urease, hydrogen peroxide, dehydrogenase, and phosphatase activities of soil, for which the degree of inhibition was closely related to its concentration [16]. Other studies found negligible effects of adding graphene to soil for the latter’s microbial structure [17] and enzyme activity [18], or that graphene reportedly caused few bacterial changes, leaving unaffected those taxa playing crucial roles in soil functioning, nor did it alter the soil microbial average well color development (AWCD) index [14].
Haplic Cambisols is the zonal soil of Northeastern China, where Changbai larch (Larix olgensis A. Henry, belonging to Pinaceae) is an important timber and afforestation tree species [19]. In this region, graphene may enter these Haplic Cambisols under L. olgensis forests in some agroforestry ecotones, thus influencing the soil’s biological properties, especially its microbial characteristics. Bacteria are a major group of organisms and the most abundantly occurring in soils. Yet how graphene may affect the biological characteristics of Haplic Cambisols, such as their resident bacteria and enzyme activities, is still unknown. Here, we applied different concentrations of graphene to Haplic Cambisols in an L. olgensis forest, and monitored the resulting changes to bacterial community structure and diversity, as well as the general soil environment (physicochemical properties and enzyme activities).
The specific objectives of this study were to: (1) investigate the effects of graphene on bacterial community diversity, physicochemical properties, and enzyme activities of Haplic Cambisols; and (2) compare the effects of different concentrations and treatment times of graphene on the bacterial community diversity and soil environments.
2. Materials and Methods
2.1. Soil Collection and Treatments
The loam A1 horizon of the Haplic Cambisols was collected from an L. olgensis forest at Maoershan Forest Research Station of Northeast Forestry University, Harbin, Heilongjiang Province, China, in September 2017. The L. olgensis plantations were growing on Typic Bori-Udic Cambisols corresponding to Haplic Cambisols (Greyic, Dystric) [20], which is zonal forest soil with high organic matter contents and well-developed horizons. Soil samples were stored in hop-pockets and taken to the laboratory. Plant residues and other impurities in soils were removed with tweezers. These samples were then powdered, sifted through 2-mm nylon screens, sufficiently mixed, and stored in black plastic bags (~500 g in each bag, 75 bags in total) at 25 °C for 3 days.
Pure graphene samples were purchased from Suzhou Tanfeng Graphene Technology Co., Ltd. (Suzhou, Jiangsu Province, China). The parameters for this graphene were as follows: color, black; concentration, 2 mg mL−1; thickness, <1 nm; sheet diameter, 0.2–10 µm; layer number, 1 or 2; specific surface area, 1000–1217 m2 g−1; purity, >99 wt %.
Soil samples were individually placed in 1000-mL beakers (~400 g, Table 1), in which they were mixed evenly with dried high-purity graphene powder to achieve different graphene contents of 0, 10, 100, and 1000 mg kg−1 (respectively expressed as A, B, C, and D). This mixing procedure went as follows: soils were placed in a sealed plastic bag, the graphene powder was added to the bag, and then the bag was repeatedly flipped until the graphene had evenly mixed with the soil. Soil water content was adjusted to 60% of maximum moisture capacity, then the soils were incubated darkly at 25 °C in an incubator at a constant temperature and humidity. Soil water content was maintained at 60% of the maximum moisture capacity by weekly adding high-purity water. The experimental layout is described in Table 2. There were five sampling and analysis times (i.e., 7, 15, 30, 60, and 90 days) after graphene treatments. For each sampling time, there were four treatments, with three beakers per treatment and 60 beakers in total. At 7, 15, 30, 60, and 90 days after graphene treatments, sampling and analysis of soils were conducted.
2.2. Soil Microbe Identifications
At 7, 15, 30, 60, and 90 days after graphene treatments, soil samples were randomly collected from each beaker (~2.0 g), immediately stored in a refrigerator at −80 °C, and taken to Shanghai Personal Biotechnology Co., Ltd. (Shanghai, China), where the 16S rDNA gene of soil bacteria was sequenced by Illumina high-throughput sequencing technology on a MiSeq platform to identify the bacterial community composition. Briefly, a 16S rDNA gene sequence analysis of each sample’s bacterial community was carried out following its amplification by PCR (polymerase chain reaction) (New England Biolabs (Beijing) LTD., Beijing, China), followed by analyses of their OTU (operational taxonomic unit) divisions, indices of alpha diversity, and beta diversity. The details of DNA extraction, 16S rDNA amplicon pyrosequencing, sequence analysis, bioinformatics, and statistical analysis are presented in Appendix A.
In current 16S rDNA gene-based microbial diversity studies, a 97% sequence similarity for the OTU threshold is usually used (roughly equivalent to the taxonomic level of sequence differences). Alpha diversity indices are often used to measure the abundance and diversity of bacterial communities in soils, which is composed of the richness index (Chao1), evenness index (ACE) and diversity indices (Simpson and Shannon). Examining beta diversity can reveal how similar bacterial community structures were found among different samples, visualized via principal coordinate analysis (PCA). The unweighted pair-group method with arithmetic means (UPGMA) and hierarchical clustering were used to observe the differences among samples. A Venn diagram was generated to visualize the shared and unique OTUs among samples, based on the occurrence of OTUs across samples/groups regardless of their relative abundance. Taxa abundances at the phylum, class, order, family, genus, and species levels were statistically compared among samples or groups by bacterial taxonomy analysis and representative sequences were taxonomically assigned using the RDP classifier. Pairwise comparisons of the sequence number (i.e., absolute abundance) of taxa at the phylum and genus levels also were carried out. Predictions of microbial metabolic function were mainly performed using a Venn diagram of common functional groups and a heatmap of bacterial community abundance for KEGG homologous gene clusters (KO), derived from a cluster analysis. Ordination plots from a redundancy analysis (RDA) were used to explore the relationships between the soil bacterial community and environmental variables.
2.3. Determination of Soil Physical and Chemical Properties
At 7, 15, 30, 60, and 90 days after graphene treatments, soil samples were randomly collected from each beaker (~200 g), air-dried, sifted through 2-mm nylon screens, mixed, and analyzed for their physical and chemical properties. The pH was determined by using the pH S-2 acidimeter (Shanghai Precision Science Instrument Co., LTD., Shanghai, China), with a 1.0 mol L−1 KCl extraction; organic matter content was determined using a TOC (total organic carbon) analyzer; hydrolytic nitrogen content was determined via the alkali reduction diffusion method; available P content was obtained through the Mo-Sb colorimetric method, with a 0.05 mol L−1 HCl–0.025 mol L−1 H2SO4 extraction; and available K content was obtained via the photometric method. All of these were assayed according to the methods of Chen [21].
2.4. Determination of Soil Enzyme Activity
Soil samples were randomly collected from each graphene treatment, air-dried, sifted through 2-mm nylon screens, mixed, and then their enzyme activities were determined. Catalase activity was measured using the methods of Johnson and Temple [22], urease activity was determined by the indophenol-blue colorimetric method [23], acid phosphatase activity was obtained using disodium phenyl phosphate colorimetry [24,25], and dehydrogenase activity was found using TTC (2,3,5-triphenyltetrazolium chloride) colorimetry [24].
2.5. Data Analysis
OriginPro v9 (OriginLab, Northampton, MA, USA) were used to make figures. Using the SPSS 18.0 software (IBM Corporation, Armonk, NY, USA), one-way analysis of variance (ANOVA), followed by Duncan’s test, was used to test for the treatment effects (graphene concentrations and incubation times) (at p < 0.05). Pearson correlations of all the parameters about soil physical and chemical properties and enzyme activities were analyzed in a randomized design with repeated measures using SPSS 18.0.
3. Results
3.1. Sequencing Results and Soil Sampling Depth Verification
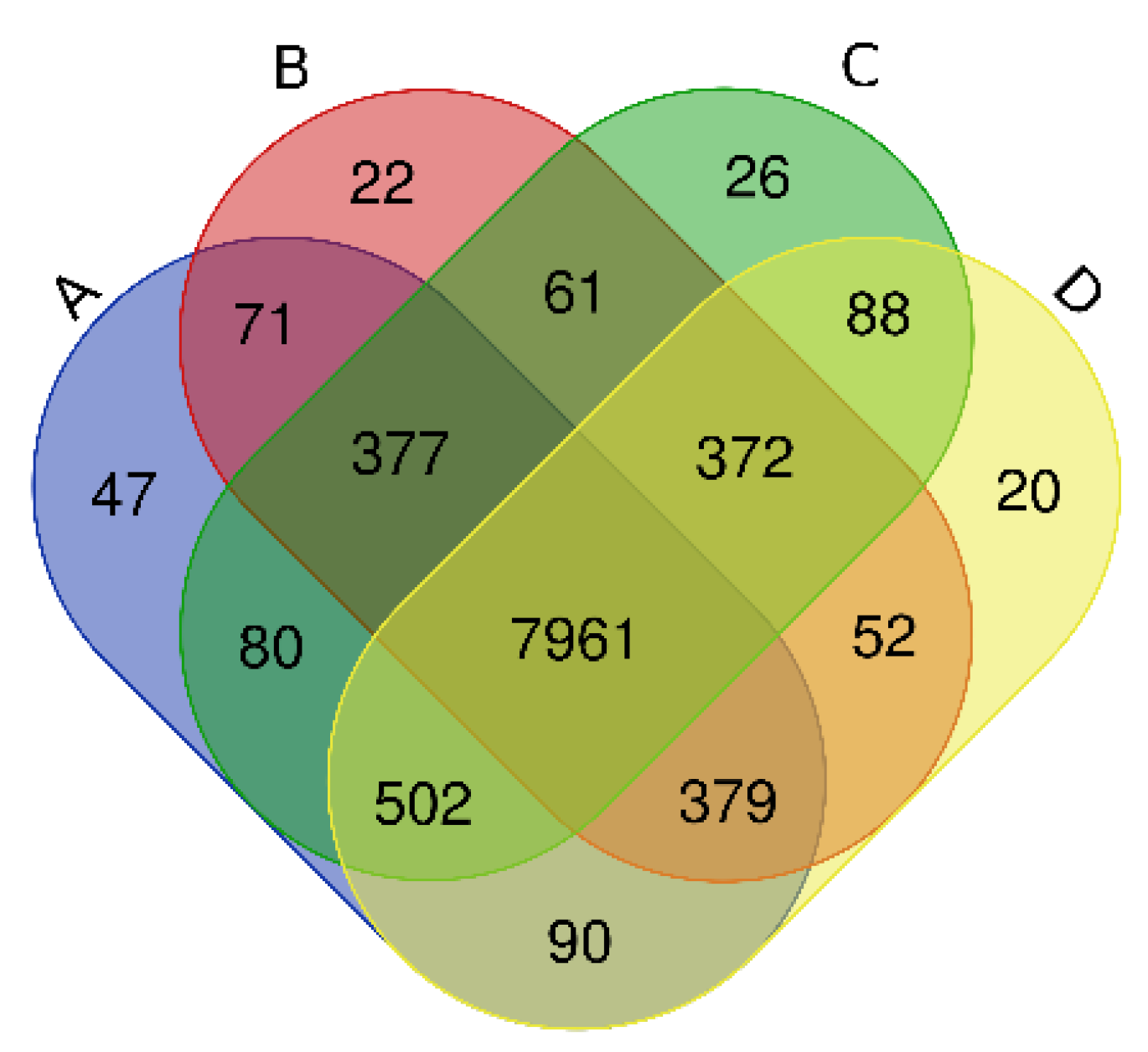
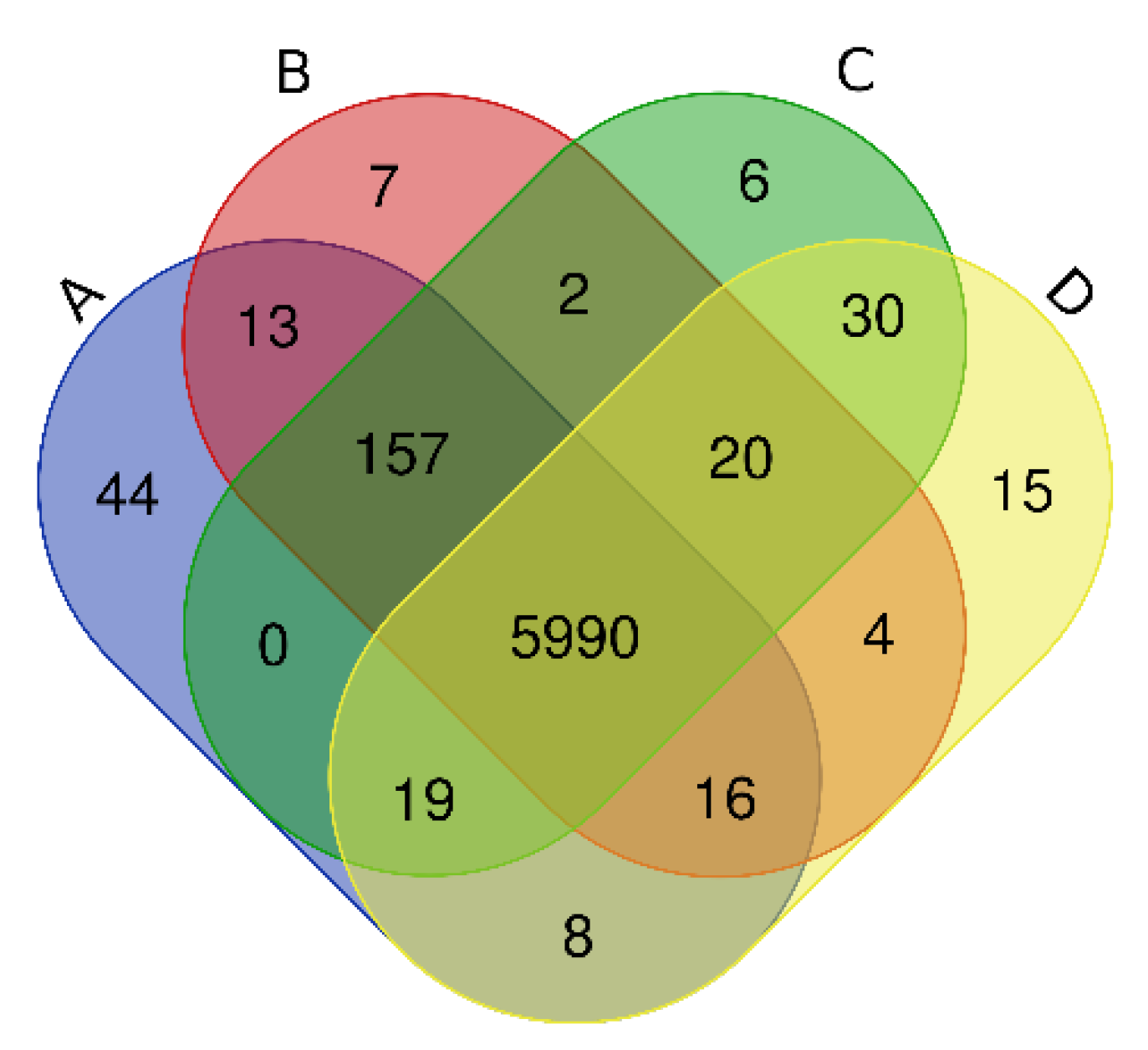
Via high-throughput sequencing, our Venn diagram (Figure 1) shows that a total of 7961 OTUs were detected in Haplic Cambisols receiving the four treatments. The OTU numbers of unique and common bacteria in 0, 10, 100, 1000 mg kg−1 (A, B, C, D) treatments were, respectively, 47, 22, 26, 20 and 9507, 9295, 9467, 9464. The number of OTUs shared by treatments B, C, and D with treatment A was 8788, 8920, and 8932, respectively.
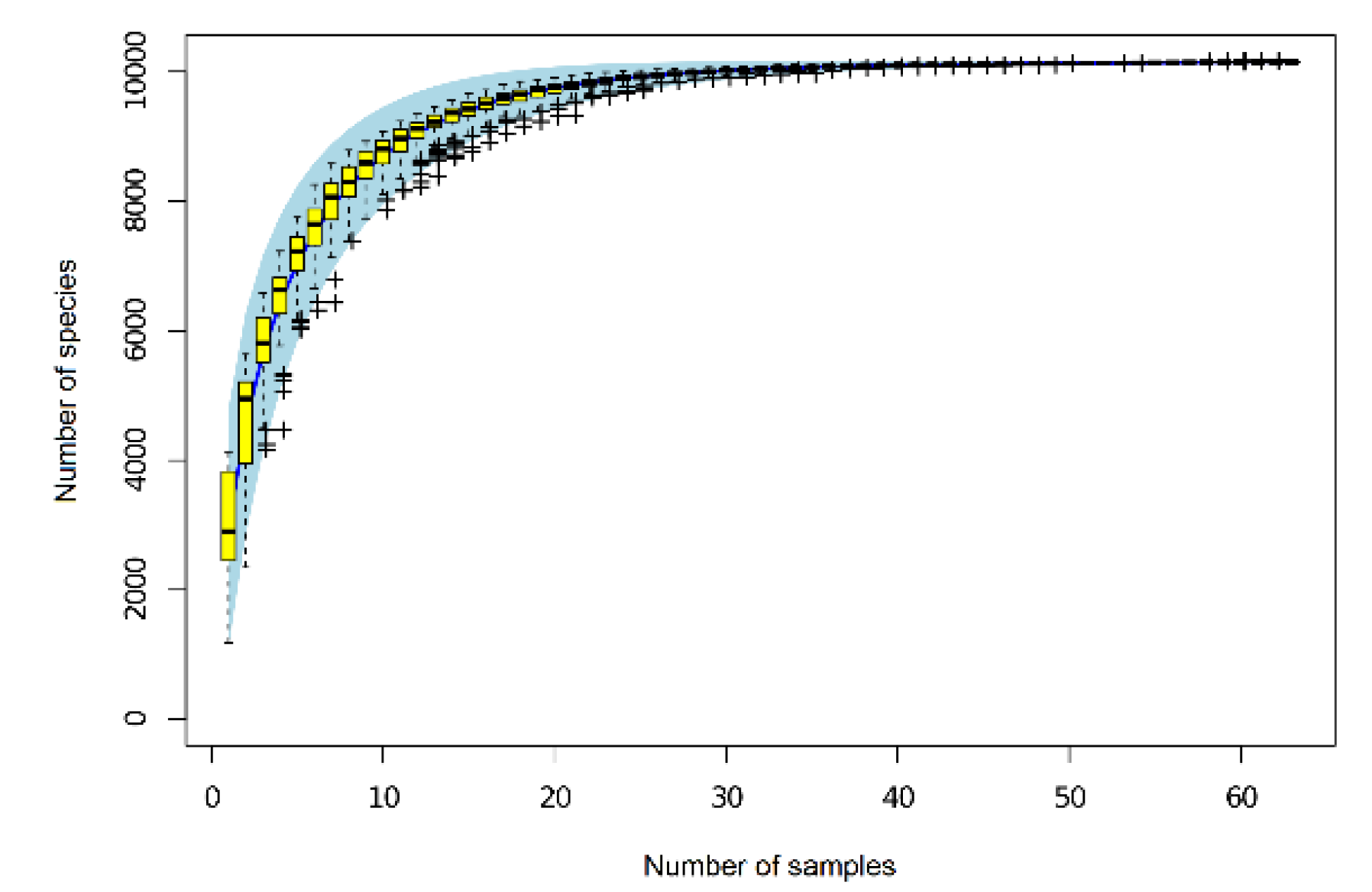
As Figure 2 shows, when the soil samples were between 30 and 40, the OTU species curve was almost flat, indicating that this sampling effort was sufficient to capture the true richness of the bacterial community. After removing the rare OTUs [26]—these amounted to <0.001% (i.e., 1/100,000) of the total sequences in all samples—the remaining OTUs covered 99.999% of the bacteria in soil. This result indicates that second-generation high-throughput Illumina MiSeq (Illumina Inc., San Diego, CA, USA) sequencing could accurately reflect the richness and diversity of soil bacteria.
3.2. Alpha Diversity Analysis
Four of the alpha diversity indices are presented in Table 3 for different graphene treatments and their durations. For species richness, the Chao1 of soil samples decreased first then increased at 7 and 15 days, indicating the number of soil OTUs was initially inhibited but rose with the graphene concentrations, reaching its maximum at 1000 mg kg−1 (D). At 30 days, the Chao1 under 0 mg kg−1 graphene (A) decreased significantly, but it increased significantly at 10 mg kg−1 (B) and 100 mg kg−1 (C), yet decreased significantly at 1000 mg kg−1 (D); this trend suggested that intermediate graphene concentrations (B and C) might promote the development of some bacteria, thus increasing soil bacteria richness, whereas if the concentration is too high it may well inhibit the growth of most bacteria, thus reducing their richness. At 60 days, the Chao1 reached its lowest values at any time, especially in the 10 mg kg−1 (B) treatment, when it was mostly similar among graphene concentrations. At 90 days, however, the Chao1 increased significantly at first and then decreased, peaking at 100 mg kg−1 (C); hence, the number of soil bacteria had increased compared with that at 60 days (lowest occurring numbers). Across the time period, this Chao1 richness index of bacteria was ranked as 15 > 7 > 30 > 90 > 60 days; the ACE index showed the same basic trend (Table 3). Both the Simpson and Shannon indices revealed that soil bacterial diversity increased slightly during the whole period in response to 10–1000 mg kg−1 of graphene (B–D). This suggested the bacterial community responded positively to more graphene input, with their associated index values rank-ordered as 15 > 7 > 90 > 30 > 60 days (Table 3).
3.3. Beta Diversity Analysis
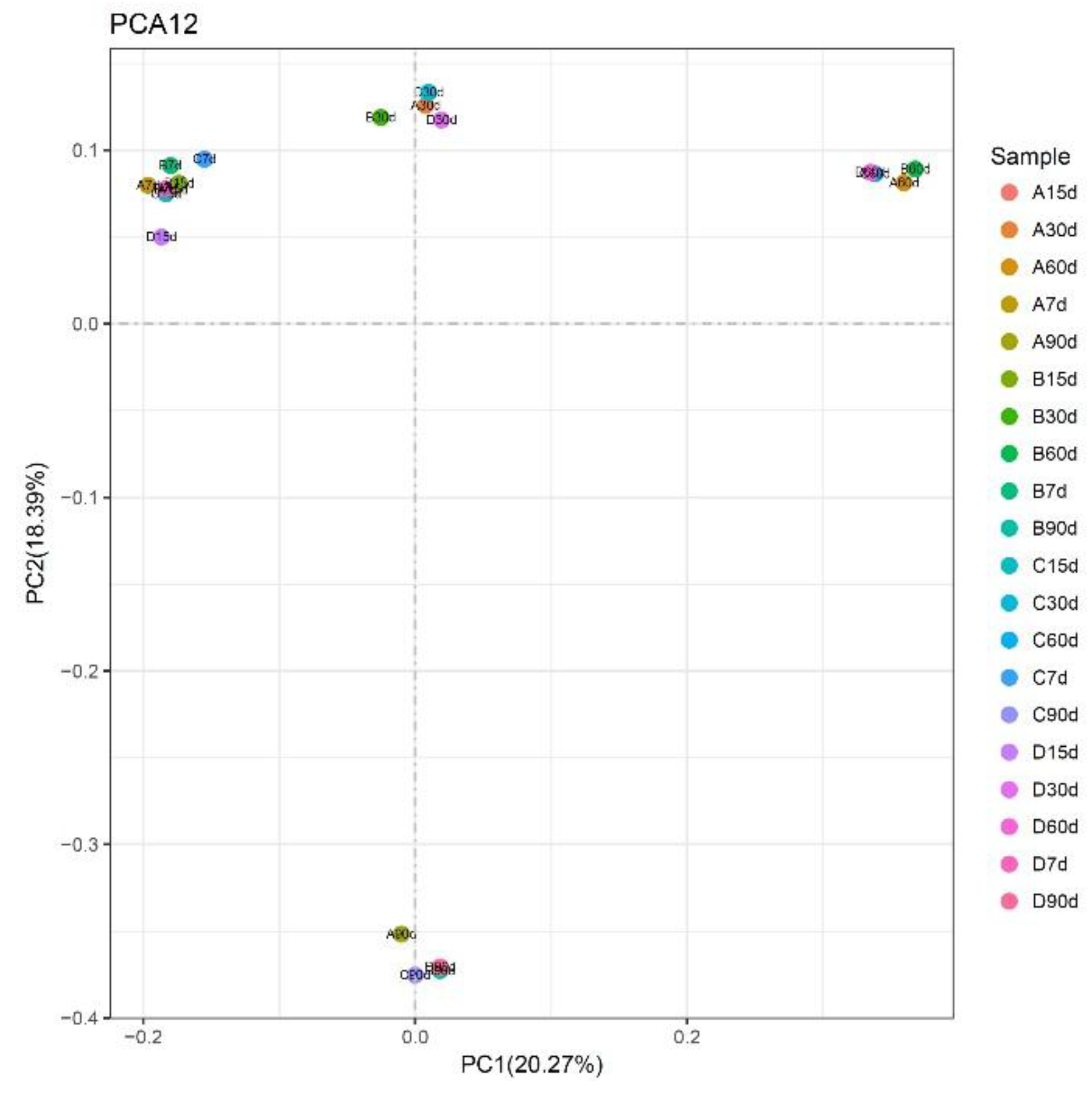
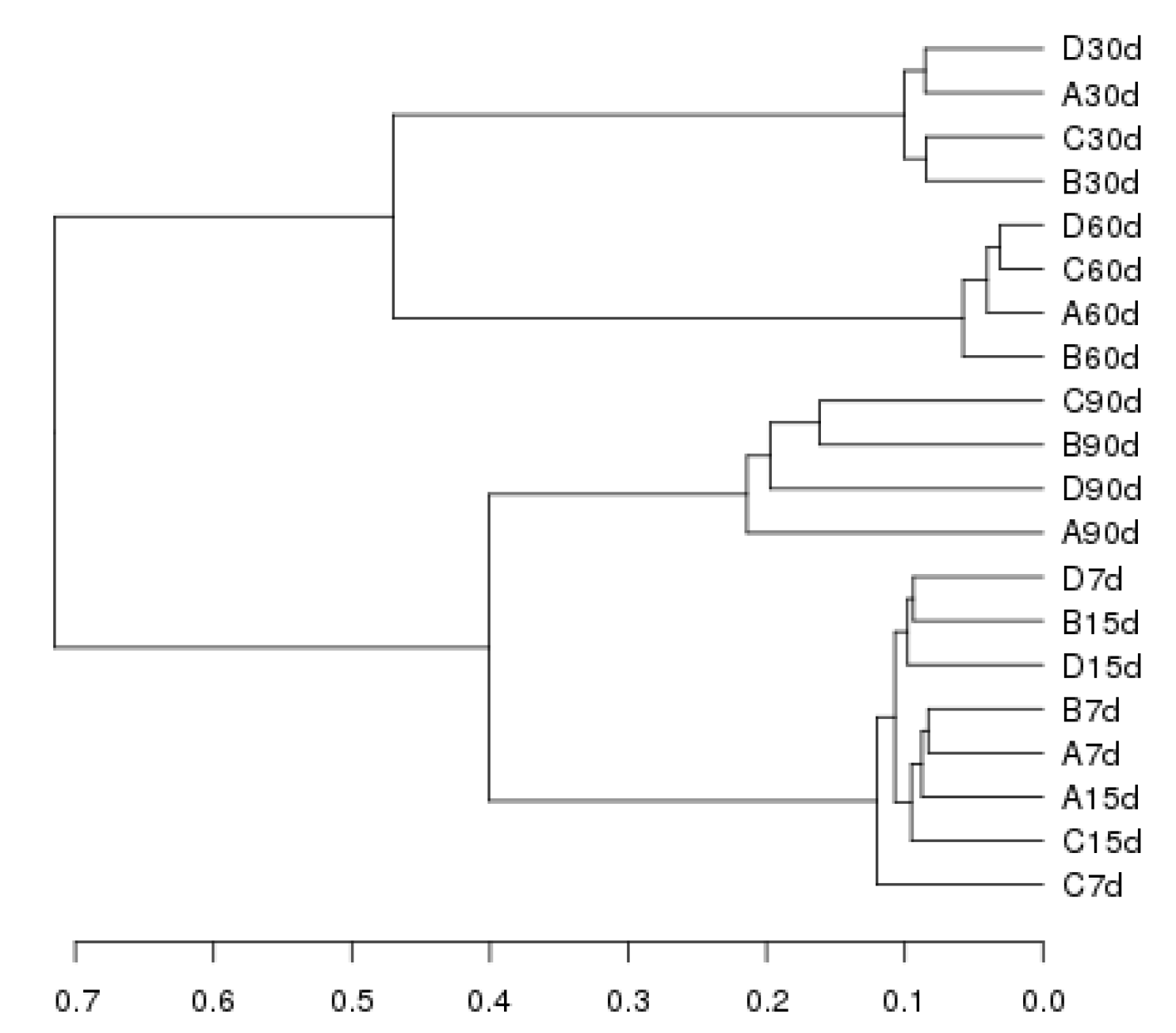
Our principal component analysis (PCA)—performed using the sample distance matrix for each soil sample sequence—showed a regular pattern in the distribution of bacterial communities under different graphene treatments (Figure 3). At 7 and 15 days, for all graphene groups (0–1000 mg kg−1; A–D), the bacteria tended to be situated to left side of the second quadrant. However, at 30 days, they had shifted their position to left side of the first quadrant, at 60 days they shifted to the right side of the first quadrant, and at 90 days they consistently occurred between the two quadrants. These results indicate that the soil bacterial community structures were relatively consistent at different incubation times. Their similarity was analyzed in a cluster analysis (Figure 4), with the ensuing order of bacterial similarity found: 15 > 7 > 30 > 60 > 90 days.
3.4. Bacterial Taxonomy Analysis
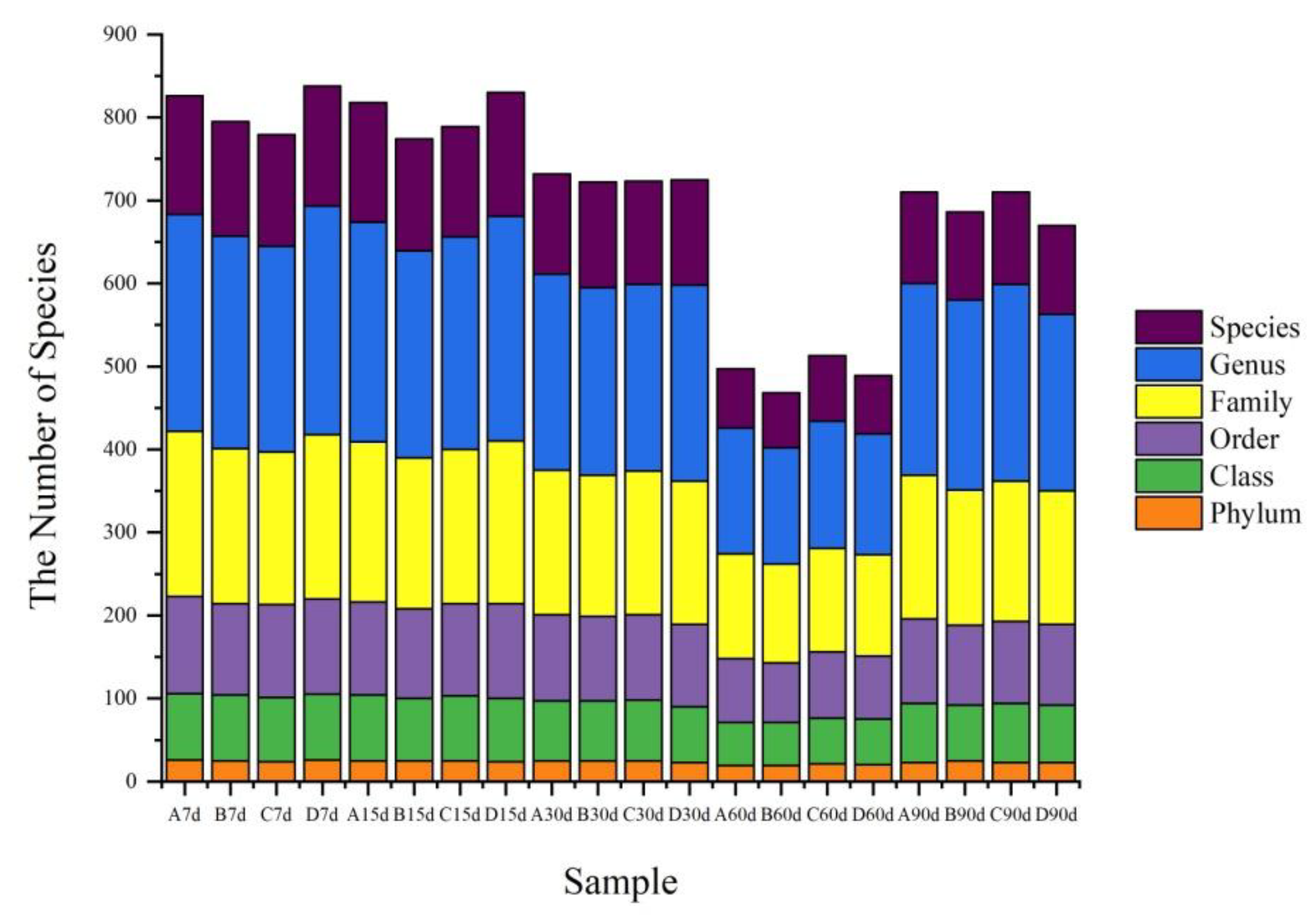
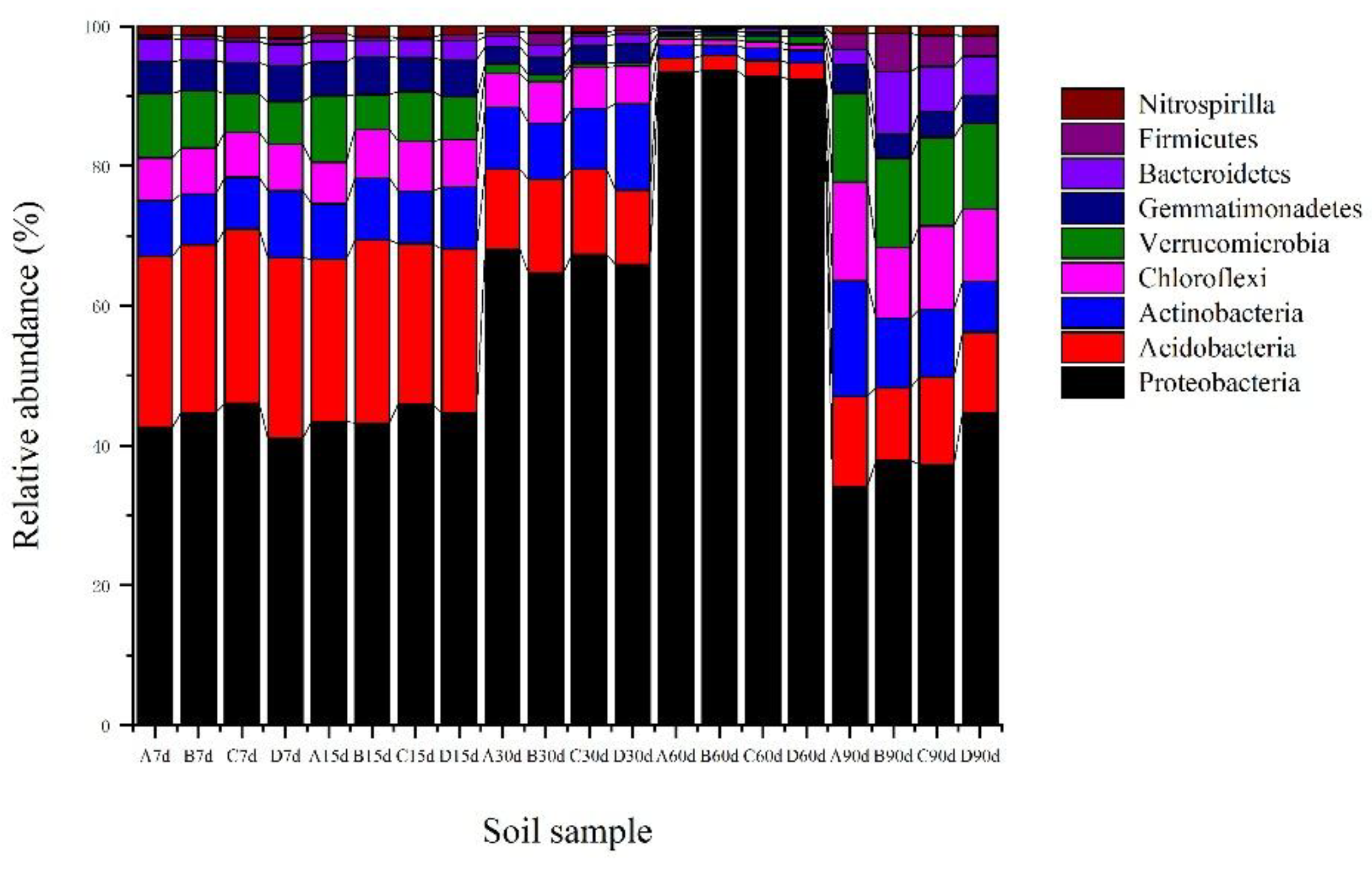
According to the population statistics (Figure 5), the number of bacterial species in soil samples varied with incubation time and graphene concentration. With graphene in the soil, a range of bacteria types were supported, including Proteobacteria, Acidobacteria, Actinobacteria, Chloroflexi, Verrucomicrobia, Gemmatimonadetes, Bacteroidetes, Firmicutes, and Nitrospirilla, as well as other bacteria (Figure 6). Except for a few unidentified bacteria, the relative abundances of Proteobacteria, Acidobacteria, Actinobacteria, Chloroflexi, and Verrucomicrobia in the soil samples exceeded those of other co-occurring taxa (Figure 7).
The relative abundance of the main bacterial community members in Haplic Cambisols varied with graphene incubation time. Compared with the 0 mg kg−1 graphene (A), Proteobacteria increased first and then decreased. The relative abundances of Acidobacteria, Actinobacteria, Chloroflexi and Verrucomicrobia all decreased initially and then increased, albeit not significantly. At 7 and 15 days, Proteobacteria increased in all treatments, though not significantly; Acidobacteria, Actinobacteria, and Chloroflexi did not change significantly in abundance, but Verrucomicrobia did decrease significantly. At 30 days, the abundances of Acidobacteria and Verrucomicrobia decreased with higher graphene concentrations while those of Proteobacteria and Actinobacteria increased; however, Chloroflexi did not change significantly. At 60 days, the abundance of Verrucomicrobia increased with more graphene, Proteobacteria decreased, yet no significant changes were detected in the other three phyla. At 90 days, the abundances of Proteobacteria and Acidobacteria both increased, while those of Actinobacteria, Chloroflexi and Verrucomicrobia decreased significantly. Throughout the incubation period, Proteobacteria and Acidobacteria were the codominant bacteria, respectively accounting for 55.4% and 14.8% of all soil bacteria in our samples (Figure 6).
We also carried out pairwise comparisons of the sequence number (i.e., absolute abundance) of taxa at the phylum and genus levels (Table 4). Their absolute abundances among the samples (groups) differed significantly among the graphene treatments. The comparisons of 0 vs. 100 mg kg−1 (A vs. C) were most different at the phylum level, while those of 0 vs. 1000 mg kg−1 (A vs. D) and 10 vs. 1000 mg kg−1 (B vs. D) had the most pronounced differences at the genus level (Table 4).
3.5. Prediction of Microbial Metabolic Function
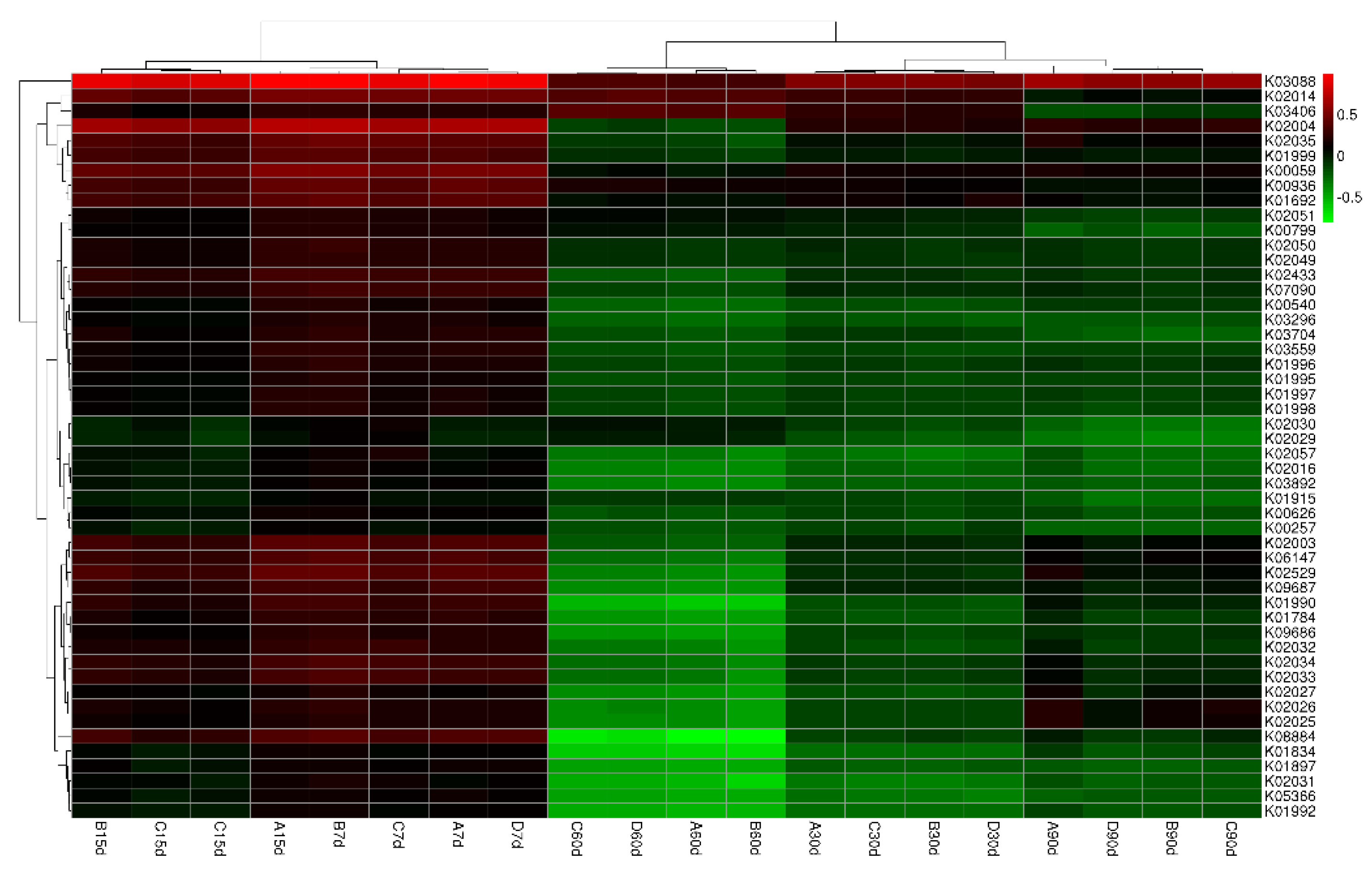
Our Venn diagram of common functional groups (Figure 8) summarized the OTU numbers of common and unique functional groups in soils. A total of 6331 OTUs were observed in the sampled Haplic Cambisols, of which 5990 were common ones (92.3%). The corresponding total numbers of OTUs for the 0, 10, 100, 1000 mg kg−1 graphene treatments (respectively A, B, C, and D) were 6247, 6209, 6224, and 6102, while their respective unique numbers were 44, 7, 6, and 15. Evidently, different concentrations of graphene significantly increased the functional groups in the bacterial community. Figure 9 shows the cluster thermal map analysis of these functional bacterial groups. Compared with 0 mg kg−1 graphene (A), the pooled abundances for 10–1000 mg kg−1 graphene (B–D) were higher at 7 days; at 15 days, those for 10–100 mg kg−1 (B–C) were higher (except for the polar amino acid transport system permease protein (K02029), the polar amino acid transport system substrate-binding protein (K02030), and the methyl-accepting chemotaxis protein (K03406)); at 30 days, the abundances of all functional bacterial groups were lower (except for the methyl-accepting chemotaxis protein (K03406) for 100–1000 mg kg−1 of graphene (C–D)); at 60 days, the abundances were likewise all lower, except for the methyl-accepting chemotaxis protein (K03406), an iron complex outer membrane receptor protein (K02014), polar amino acid transport system permease protein (K02029), and a polar amino acid transport system substrate-binding protein (K02030) for 100–1000 mg kg−1 graphene (C–D); at 90 days, the abundances were still lower, except that the abundances of multiple sugar transport system substrate-binding protein (K02027) and multiple sugar transport system permease protein (K02026) were higher in the 0 mg kg−1 (A) and 100 mg kg−1 (C) graphene treatments.
3.6. Changes in the Soil Environment
From the correlations between soil physical and chemical properties and enzyme activities (Table 5), soil pH and hydrolytic nitrogen contents were evidently strongly and positively associated, as were available potassium contents and urease activities (p < 0.01). Notable positive correlations (p < 0.01), noteworthy negative correlations (p < 0.01), and significant positive correlations (p < 0.05) also occurred between some parameters. The many correlations found, for which some were much stronger than others, suggested that ecological or environmental information overlapped among them.
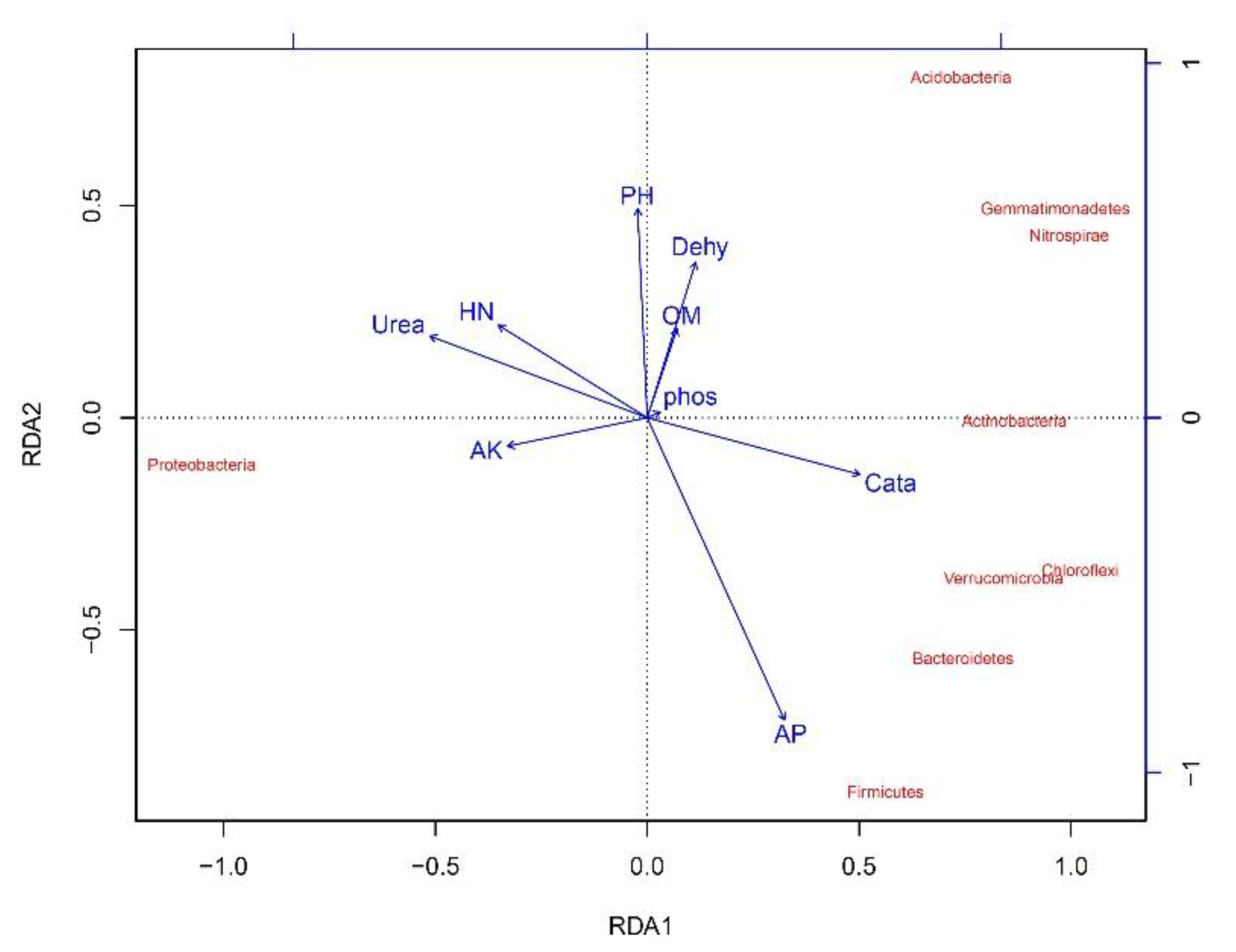
From the redundancy analysis (RDA) (Figure 10), some soil environmental factors, such as organic matter and hydrolytic nitrogen contents, as well as urease and catalase activities, also heavily influenced the soil bacterial community diversity in Haplic Cambisols. Positive correlations were found between hydrolytic nitrogen contents and urease activities and Proteobacteria. Positive correlations were also found between Acidobacteria, Actinobacteria, Chloroflexi, Verrucomicrobia, Gemmatimonadetes and organic matter contents and catalase activities. However, negative correlations occurred between Acidobacteria, Actinobacteria, Chloroflexi, Verrucomicrobia, Gemmatimonadetes and hydrolytic nitrogen contents and urease activities.
4. Discussion
After incubating the Haplic Cambisols with different concentrations of graphene, their bacterial community structure, richness, and diversity, as well as the relative abundance of their main bacterial populations, were changed (Table 3). Early in the graphene treatment, bacterial abundance decreased but then increased under greater graphene concentrations (Table 3), which suggested that microorganisms responded rapidly to environmental changes due to their high surface-to-volume ratios and low homeostasis ability [28]. Under pure incubation conditions, the contact of graphene with bacterial surfaces damages their cell membrane, thereby reducing their survival, which is consistent with the findings of Akhavan and Ghaderi [15]. With more graphene, the numbers of bacteria in Haplic Cambisols increased, indicating that higher graphene concentrations (i.e., 100 and 1000 mg kg−1) promoted bacterial growth, unlike the results reported by Ren et al. [29]. With prolonged graphene incubation times, under low concentrations of graphene, bacterial abundances increased; however, at high concentrations, they decreased significantly. This result suggests that low concentrations of graphene may promote soil bacteria to a certain extent, thus increasing their abundances and number of species. However, high graphene concentrations may inhibit the growth of some bacteria and significantly reduce their abundances in the community. For example, the lowest bacterial abundance occurred at 60 days and under 10 mg kg−1 graphene (B).
PCA (Figure 3) and cluster (Figure 4) analyses can elucidate potential changes in the structure of soil bacterial communities exposed to graphene. As Figure 3 shows, bacteria strongly overlapped between samples early on in the graphene treatment (at 7 and 15 days), when only the high concentration of graphene (100 mg kg−1, i.e., C) was distinguishable from the other treatments, indicating that graphene had little impact on bacteria at these times. At 30 days, however, graphene greatly changed the structure of the bacterial community in Haplic Cambisols, with significant differences evident among bacterial community samples. At 60 days, the differences in the bacterial community among the samples were greatest, and a high graphene concentration (100 and 1000 mg kg−1, i.e., C and D) was crucial for promoting the community-level transformation. At 90 days, the bacterial communities of all treatments tended to be consistent. The cluster analysis showed that the bacterial community increasingly diverged as the incubation time lengthened, and these differences were most pronounced at 60 days (Figure 4). Further, although the dominant community composition at the phylum level was similar among the different graphene treatments, their relative abundances varied (Figure 6). In the five graphene treatment communities, the abundance order for Proteobacteria was 60 > 30 > 7 > 15 > 90 days, while that for Acidobacteria and Actinobacteria was 7 > 15 > 90 > 30 > 60 days, corresponding to graphene’s impact on microorganisms at 60 days, which led to predictable changes in soil physicochemical properties and microbial selection. The abundances of Acidobacteria and Actinobacteria were higher in early than late stages of incubation, likely because both can degrade cellulose (even refractory aromatic compounds) and promote soil mineralization and nitrogen cycling.
Proteobacteria, Acidobacteria, Actinobacteria, Chloroflexi, and Verrucomicrobia were variously dominant soil microorganisms in soil with different graphene and incubation times (Figure 6 and Figure 7). Buckley and Schmidt [30] as well as Tang et al. [31] found that Proteobacteria, Acidobacteria, Planctomycetes, Actinobacteria, and Bacteroidetes are the dominant phyla in agricultural soils. In our study, three of these were among our five dominant phyla, which indicated similarities and differences in soil microorganisms between Haplic Cambisols and agricultural soils. This interpretation is consistent with the considerable community differences we found at the phylum and genus levels among the graphene treatments (Table 4).
Changes in the quantity of soil microbes may be used to gauge the effect of graphene on soil ecosystem stability [32]. We found that changes to main bacterial community members were closely related to graphene concentration and incubation time (Figure 8). Soon after the graphene additions, the metabolic functions of bacterial communities remained unchanged. However, with the 1000 mg kg−1 graphene (D) treatment, the soil bacteria numbers clearly increased. Early on in the incubation, 10–1000 mg kg−1 graphene (B–D) affected the structure and function of the soil bacterial community. The genes of a carrier protein and a substrate binding enzyme protein related to amino acid transport to the cell membrane of bacteria, in addition to methyl-accepting chemotaxis signal transduction protein (MCP), all had decreased, which should affect the preparation of the MCP recombinant protein. With a longer incubation time, the metabolic function of the soil bacterial community decreased continuously, but the MCP gene began to increase, not yet binding to graphene, but to the cholinesterase A (CheA) phosphorylated by ATP. This phosphorylated protein may transfer its phosphoester bond to the cholinesterase Y protein, and the interaction between the Y protein and FliM switching protein in the flagellum matrix promoted the flagellum rotation and cell roll of Pseudomonas, which reached its maximum at 60 days, followed by 30 days (Figure 8). Though bacterial communities can resist graphene treatment and promote cell growth and reproduction through a series of reactions (for example, a protein-iron can combine with its receptor and form a complex in the cytoplasm or interstitial fluid, participating in DNA synthesis in the cell as induced by the increase of carrier proteins and substrate-binding enzymes on the cell membrane of the bacteria), the effect of such reactions is likely very small. This would explain why the bacterial community richness and diversity were lowest at 60 days. At 90 days, the multiple sugar transport system substrate-binding protein and multiple sugar transport system permease protein were more active in their roles under 100 mg kg−1 graphene (C), with lipopolysaccharide transporters (LptA-G) initiating transmembrane transport (lipopolysaccharides are essential for the survival of all Gram-negative bacteria). Permease proteins (carrier proteins), being a kind of membrane protein, can improve the cell membrane permeability to specific molecules, and can bind to extracellular substances and then release these substances via the membrane for cell growth in a process that does not consume metabolic energy. Hence, incubation time and graphene concentration both drive the bacterial community’s metabolic functioning (Figure 9).
Changes in the bacterial community in Haplic Cambisols were also related to soil physical and chemical properties and enzyme activities. Our trends for soil bacterial community richness and diversity indices were consistent with soil organic matter and hydrolytic nitrogen contents, as well as urease activities, among others. This is perhaps not surprising, since soil organic matter is a key nutrient and energy source of microorganisms [24], and urease can specifically promote urea hydrolysis to ammonia and carbonic acid, and is an important source of microbial nitrogen sources [33,34]. Graphene likely affected the nutrient and energy sources of soil bacteria by changing the organic matter contents available to them. Hydrolytic nitrogen and urease are key sources of bacterial nitrogen; thus, graphene will likely increase soil nitrogen cycling in these Haplic Cambisols, with hydrolytic nitrogen contents and urease activities significantly increased in the soil bacterial community at 7 and 15 days (Figure 10). Soil catalase activities are related to the numbers of species and the activity of the bacterial community, which can reflect the intensity of the soil microbiological process to a certain extent. In our study, only early in graphene incubation did the catalase activities of soils notably increase (Figure 10), thus indicating that the direct contact of graphene with catalase probably changed the enzyme structure and properties, with soil bacteria producing reactive oxygen species (ROS). This view is consistent with the findings of Ahmed and Rodrigues [35] and Wang et al. [18].
With a longer incubation time, catalase activities were mostly unchanged, indicating that the effects of graphene on soils were limited, as found by Li et al. [14]. Dehydrogenase activity is both related to microbial respiration and biomass, thus it could indirectly reflect the activity and viability of microorganisms [36]. In this study, the effect of graphene on soil dehydrogenase was relatively small: only at 15 and 30 days did its activity exceed that under 0 mg kg−1 graphene (A). Compared with the 0 mg kg−1 graphene treatment (A), the activity of acid phosphatase increased significantly at the early stage of graphene incubation, and decreased significantly at 60 and 90 days, indicating that graphene may affect soil N and P cycling when it reaches a certain content in the soil (Table 5; Figure 10).
5. Conclusions
Graphene increased the richness and diversity indices of the bacteria community in Haplic Cambisols, particularly during early stages of incubation (7 and 15 days). The order of bacterial similarity in Haplic Cambisols treated with graphene was 15 > 7 > 30 > 60 > 90 days, i.e., only at later stages did graphene significantly increase the bacterial community structure (i.e., after 30 days). The relative abundances of Proteobacteria and Acidobacteria in Haplic Cambisols were highest, with the number of Pseudomonas of Proteobacteria being particularly large. The specie numbers of bacteria in Haplic Cambisols varied with graphene concentration, but more strongly with incubation time; its rank order was 15 > 7 > 30 > 90 > 60 days.
Soil environmental factors, such as organic matter and hydrolytic nitrogen contents as well as urease and catalase activities, also influenced the soil bacterial community diversity in Haplic Cambisols. Since their trends closely matched those found for bacterial richness and diversity, these environmental factors are likely linked to processes of bacterial community assembly in Haplic Cambisols.
Author Contributions
Conceptualization, J.S. and X.C.; methodology, J.S., C.D., and X.C.; investigation, C.D., S.W., and J.R.; data curation, C.D. S.W., and J.R.; writing—original draft preparation, C.D., Y.S., and J.S.; writing—review and editing, C.D., Y.S., J.S., and X.C.; project administration, J.S.
Funding
This work was financially supported by the National Natural Science Foundation of China (31370613), the Fundamental Research Funds for the Central Universities (2572017CA03), and the Harbin Science and Technology Bureau Youth Science and Technology Innovation Project (2102RFQXN019).
Acknowledgments
We thank Shanghai Personal Biotechnology Co., Ltd. for the identifications of bacterial community composition. We are grateful to the editors and reviewers for their help and valuable suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A. Sequencing of 16S rDNA Gene in Soil Bacteria
Appendix A.1. DNA Extraction
Total bacterial genomic DNA samples were extracted using the Fast DNA SPIN extraction kits (MP Biomedicals, Santa Ana, CA, USA), and stored at −20 °C prior to further analysis. The quantity and quality of extracted DNAs were measured using a NanoDrop ND-1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and agarose gel electrophoresis, respectively.
Appendix A.2. 16S rDNA Amplicon Pyrosequencing
PCR amplification of the bacterial 16S rDNA genes V3–V4 region was performed using the forward primer 338F (5’-ACTCCTACGGGAGGCAGCA-3’) and the reverse primer 806R (5’-GGACTACHVGGGTWTCTAAT-3’). Sample-specific 7-bp barcodes were incorporated into the primers for multiplex sequencing. The PCR components contained 5 μL of Q5 reaction buffer (5×), 5 μL of Q5 High-Fidelity GC buffer (5×), 0.25 μL of Q5 High-Fidelity DNA Polymerase (5U/μL), 2 μL (2.5 mM) of dNTPs, 1 μL (10 μM) of each Forward and Reverse primer, 2 μL of DNA Template, and 8.75 μL of ddH2O. Thermal cycling consisted of initial denaturation at 98 °C for 2 min, followed by 25 cycles consisting of denaturation at 98 °C for 15 s, annealing at 55 °C for 30 s, and extension at 72 °C for 30 s, with a final extension of 5 min at 72 °C. PCR amplicons were purified with Agencourt AMPure Beads (Beckman Coulter, Indianapolis, IN, USA) and quantified using the PicoGreen dsDNA Assay Kit (Invitrogen, Carlsbad, CA, USA). After the individual quantification step, amplicons were pooled in equal amounts, and pair-end 2 × 300 bp sequencing was performed using the Illlumina MiSeq (Illumina Inc., San Diego, CA, USA) platform with MiSeq Reagent Kit v3 at Shanghai Personal Biotechnology Co., Ltd (Shanghai, China).
Appendix A.3. Sequence Analysis
The Quantitative Insights Into Microbial Ecology (QIIME, v1.8.0) pipeline was employed to process the sequencing data. Briefly, raw sequencing reads with exact matches to the barcodes were assigned to respective samples and identified as valid sequences. The low-quality sequences were filtered through following criteria [37]: sequences that had a length of <150 bp, sequences that had average Phred scores of <20, sequences that contained ambiguous bases, and sequences that contained mononucleotide repeats of >8 bp. Paired-end reads were assembled using FLASH [38]. After chimera detection, the remaining high-quality sequences were clustered into operational taxonomic units (OTUs) at 97% sequence identity by UCLUST [39]. A representative sequence was selected from each OTU using default parameters. An OTU table was further generated to record the abundance of each OTU in each sample and the taxonomy of these OTUs. OTUs containing less than 0.001% of total sequences across all samples were discarded. To minimize the difference of sequencing depth across samples, an averaged, rounded rarefied OTU table was generated by averaging 100 evenly resampled OTU subsets under the 90% of the minimum sequencing depth for further analysis.
Appendix A.4. Bioinformatics and Statistical Analysis
Sequence data analyses were mainly performed using QIIME and R packages (v3.2.0). OTU-level alpha diversity indices, such as richness index (Chao1), evenness index (ACE) and diversity indices (Simpson and Shannon), were calculated using the OTU table in QIIME. OTU-level ranked abundance curves were generated to compare the richness and evenness of OTUs among samples. Beta diversity analysis was performed to investigate the structural variation of microbial communities across samples using UniFrac distance metrics [40], visualized via principal coordinate analysis (PCA), and unweighted pair-group method with arithmetic means (UPGMA) hierarchical clustering [41]. Venn diagram was generated to visualize the shared and unique OTUs among samples or groups using R package “VennDiagram”, based on the occurrence of OTUs across samples/groups regardless of their relative abundance [42]. Taxa abundances at the phylum, class, order, family, genus and species levels were statistically compared among samples or groups by bacterial taxonomy analysis and representative sequences were taxonomically assigned using the RDP classifier. Pairwise comparisons of sequence number (i.e., absolute abundance) of taxa at the phylum and genus levels also were carried out. Prediction of microbial metabolic function were mainly performed using Venn diagram of common functional groups and Heatmap of bacterial community abundance for KEGG homologous gene clusters (KO), derived from a cluster analysis. Ordination plots from a redundancy analysis (RDA) used to explore the relationships between the soil bacterial community and environmental variables.
References
- Robinson, J.T.; Tabakman, S.M.; Liang, Y.; Wang, H.; Casalongue, H.S. Ultrasmall reduced graphene oxide with high near-infrared absorbance for photothermal therapy. J. Am. Chem. Soc. 2011, 133, 6825–6831. [Google Scholar] [CrossRef] [PubMed]
- Novoselov, K.S.; Geim, A.K.; Morozov, S.V.; Jiang, D.; Zhang, Y.; Dubonos, S.V.; Grigorieva, I.V.; Firsov, A.A. Electric field effect in atomically thin carbon films. Science 2004, 306, 666–669. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Jang, C.; Xiao, S.D.; Ishigami, M.; Fuhrer, M.S. Intrinsic and extrinsic performance limits of graphene devices on SiO2. Nat. Nanotechnol. 2008, 3, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Service, R.F. Materials science. Carbon sheets an atom thick give rise to graphene dreams. Science 2009, 324, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Wei, X.; Kysar, J.W.; Hone, J. Measurement of the elastic properties and intrinsic strength of monolayer graphene. Science 2008, 321, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.J.; Qin, J.G.; Li, Z. Research advances of graphene. Prog. Chem. 2016, 35, 2559–2567. [Google Scholar]
- Klaine, S.J.; Alvarez, P.J.J.; Batley, G.E.; Lead, J.R. Critical review—Nanomaterials in the environment: Behavior, fate, bioavailability, and effects. Environ. Toxicol. Chem. 2008, 27, 1825–1851. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Yu, F.; Ma, J.; Chen, J.H. Research progress of application of graphene on removing heavy metals and antibiotics from water. J. Funct. Mater. 2014, 23, 23001–23009. [Google Scholar]
- Singh, N.; Srivastava, G.; Talat, M.; Raghubanshi, H.; Srivastava, O.N.; Kayastha, A.M. Cicer, α-galactosidase immobilization onto functionalized graphene nanosheets using response surface method and its applications. Food Chem. 2014, 142, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.M.; Song, Z.L.; Parr, J.F.; Wang, H.L. Occluded C in rice phytoliths: Implications to biogeochemical carbon sequestration. Plant Soil 2013, 370, 615–623. [Google Scholar] [CrossRef]
- Fan, H.L.; Xie, A.Q.; Shen, C.; Su, S.; Wu, C.Z.; Hong, W. Isolation, identification and oxidant activity of endophytic fungi from Liriopemuscari (Decne.) Bailey. J. Beihua Univ. (Nat. Sci.) 2017, 18, 106–109. [Google Scholar]
- Yan, J.; Han, X.Z.; Ji, Z.J.; Li, Y.; Wang, E.T.; Xie, Z.H.; Chen, W.F. Abundance and diversity of soybean-nodulating rhizobia in black soil are impacted by land use and crop management. Appl. Environ. Microbiol. 2014, 80, 5394–5402. [Google Scholar] [CrossRef] [PubMed]
- Badiane, N.N.Y.; Chotte, J.L.; Pate, E.; Masse, D.; Rouland, C. Use of soil enzyme activities to monitor soil quality in natural and improved fallows in semi-arid tropical regions. Appl. Soil Ecol. 2001, 18, 229–238. [Google Scholar] [CrossRef]
- Li, L.N.; Teng, Y.; Ren, W.J.; Li, Z.G.; Luo, Y.M. Effects of Graphene on soil enzyme activities and microbial communities. Soil 2016, 48, 102–108. [Google Scholar]
- Akhavan, O.; Ghaderi, E. Toxicity of graphene and graphene oxide nanowalls against bacteria. ACS Nano 2010, 4, 5731–5736. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J. The Biological Effect of Nanocapsule Pesticide and Carbon Nanomaterials on Soil Microorganism. Ph.D. Thesis, University of Science and Technology Beijing, Beijing, China, 2015. [Google Scholar]
- Karmali, P.P.; Simberg, D. Interactions of nanoparticles with plasma proteins: Implication on clearance and toxicity of drug delivery systems. Expert Opin. Drug Deliv. 2011, 8, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.S.; Qi, X.C.; Shen, T.L.; Li, C.L. Effect of silver nanoparticles and graphene on soil microorganisms and enzyme activities. Acta Sci. Circumst. 2017, 37, 3149–3157. [Google Scholar]
- Yang, J.; He, Y.; Aubrey, D.P.; Zhuang, Q.; Teskey, R.O. Global patterns and predictors of stem CO2 efflux in forest ecosystems. Glob. Chang. Biol. 2016, 22, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2006; World Soil Resource Reports 103; FAO: Rome, Italy, 2006; pp. 53–65. [Google Scholar]
- Chen, L.X. Soil Experiment and Practice Course; Northeast Forestry University Press: Harbin, China, 2005; pp. 61–95. (In Chinese) [Google Scholar]
- Johnson, J.L.; Temple, K.L. Some variables affecting the measurement of “catalase activity” in soil. Soil Sci. Soc. Am. J. 1964, 28, 207–209. [Google Scholar] [CrossRef]
- Hoffmann, G.; Teicher, K. Ein kolorimetrisches verfahren zur bestimmung der ureaseaktivität in Böden. J. Plant Nutr. Soil Sci. 2010, 95, 55–63. [Google Scholar] [CrossRef]
- Guan, S.Y. Soil Enzyme and Its Research Method; Agricultural Press: Beijing, China, 1986; pp. 274–339. (In Chinese) [Google Scholar]
- Zheng, H.Y.; Zhang, D.S.T. Research Methods of Soil Dynamic Biochemistry; Science Press: Beijing, China, 1982; pp. 53–56. (In Chinese) [Google Scholar]
- Bokulich, N.A.; Collins, T.S.; Chad, M.; Greg, A.; Hildegarde, H.; Ebeler, S.E.; Mills, D.A. Associations among wine grape microbiome, metabolome, and fermentation behavior suggest microbial contribution to regional wine characteristics. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Asnicar, F.; Weingart, G.; Tickle, T.L.; Huttenhower, C.; Segata, N. Compact graphical representation of phylogenetic data and metadata with GraPhlAn. PeerJ 2015, 3, e1029. [Google Scholar] [CrossRef] [PubMed]
- Boivin, M.E.Y.; Breure, A.M.; Posthuma, L.; Rutgers, M. Determination of field effects of contaminants– significance of pollution-induced community tolerance, Hum. Ecol. Risk Assess. 2002, 8, 1035–1055. [Google Scholar] [CrossRef]
- Ren, W.; Ren, G.; Teng, Y.; Li, Z.; Li, L. Time-dependent effect of graphene on the structure, abundance, and function of the soil bacterial community. J. Hazard. Mater. 2015, 297, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Buckley, D.H.; Schmidt, T.M. Diversity and dynamics of microbial communities in soils from agro- ecosystems. Environ. Microbiol. 2003, 5, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Yuan, B.; Lai, Q.; Wang, R.; Bao, H.; Feng, F.Y. Hymenobacter terrenus sp. nov., isolated from biological soil crusts. Int. J. Syst. Evol. Microbiol. 2015, 65, 4557–4562. [Google Scholar] [CrossRef] [PubMed]
- Doran, J.W.; Zeiss, M.R. Soil health and sustainability: Managing the biotic component of soil quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef]
- Fu, L.F.; Li, M.L.; Qin, H.M.; Yin, H.; Mo, C.H. Effects of decabromodiphenyl ether on enzyme activities of two type soils. Soil 2014, 46, 689–696. [Google Scholar]
- Zhang, C.; Chen, X.F.; Zhou, B.; Zhang, C.L.; Li, J.J.; Zhang, J.; Dai, J. Effects of vermicompost on microbial characteristics and enzyme activities in soil. Soils 2014, 46, 70–75. [Google Scholar]
- Ahmed, F.; Rodrigues, D.F. Investigation of acute effects of graphene oxide on wastewater microbial community: A case study. J. Hazard. Mater. 2013, 256, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Neto, M.; Ohannessian, A.; Delolme, C.; Bedell, J.P. Towards an optimized protocol for measuring global dehydrogenase activity in storm-water sediments. J. Soil Sediments 2007, 7, 101–110. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, W. Application of high-throughput sequencing in understanding human oral microbiome related with health and disease. Front. Microbiol. 2014, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microb. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Ramette, A. Multivariate analyses in microbial ecology. Fems Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaura, E.; Keijser, B.J.F.; Huse, S.M.; Crielaard, W. Defining the healthy “core microbiome” of oral microbial communities. Bmc Microbiol. 2009, 9, 12. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Venn diagrams of the total operational taxonomic units (OTUs) in Haplic Cambisols treated with different concentrations of graphene and different incubation times (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1).
Figure 1.
Venn diagrams of the total operational taxonomic units (OTUs) in Haplic Cambisols treated with different concentrations of graphene and different incubation times (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1).

Figure 2.
Species accumulation curves (boxplots) in Haplic Cambisols treated with graphene.

Figure 3.
Principal component analysis (PCA) analysis of soil bacterial communities under different graphene treatments (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1; 7, 15, 30, 60, and 90 d mean 7, 15, 30, 60, and 90 days, respectively).
Figure 3.
Principal component analysis (PCA) analysis of soil bacterial communities under different graphene treatments (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1; 7, 15, 30, 60, and 90 d mean 7, 15, 30, 60, and 90 days, respectively).

Figure 4.
An unweighted, pair-group method with arithmetic means (UPGMA) clustering analysis chart, based on a weighted UniFrac distance matrix. Soil samples are clustered according to their similarity: the shorter the branch length between two samples, the greater their similarity. (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1; 7, 15, 30, 60, and 90 d mean 7, 15, 30, 60, and 90 days, respectively).
Figure 4.
An unweighted, pair-group method with arithmetic means (UPGMA) clustering analysis chart, based on a weighted UniFrac distance matrix. Soil samples are clustered according to their similarity: the shorter the branch length between two samples, the greater their similarity. (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1; 7, 15, 30, 60, and 90 d mean 7, 15, 30, 60, and 90 days, respectively).

Figure 5.
Statistics of the soil bacterial groups at different taxonomic levels. (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1; 7, 15, 30, 60, and 90 d mean 7, 15, 30, 60, and 90 days, respectively).
Figure 5.
Statistics of the soil bacterial groups at different taxonomic levels. (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1; 7, 15, 30, 60, and 90 d mean 7, 15, 30, 60, and 90 days, respectively).

Figure 6.
Bacterial compositions of the nine most abundant phyla in the soil samples treated with graphene (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1; 7, 15, 30, 60, and 90 d mean 7, 15, 30, 60, and 90 days, respectively).
Figure 6.
Bacterial compositions of the nine most abundant phyla in the soil samples treated with graphene (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1; 7, 15, 30, 60, and 90 d mean 7, 15, 30, 60, and 90 days, respectively).

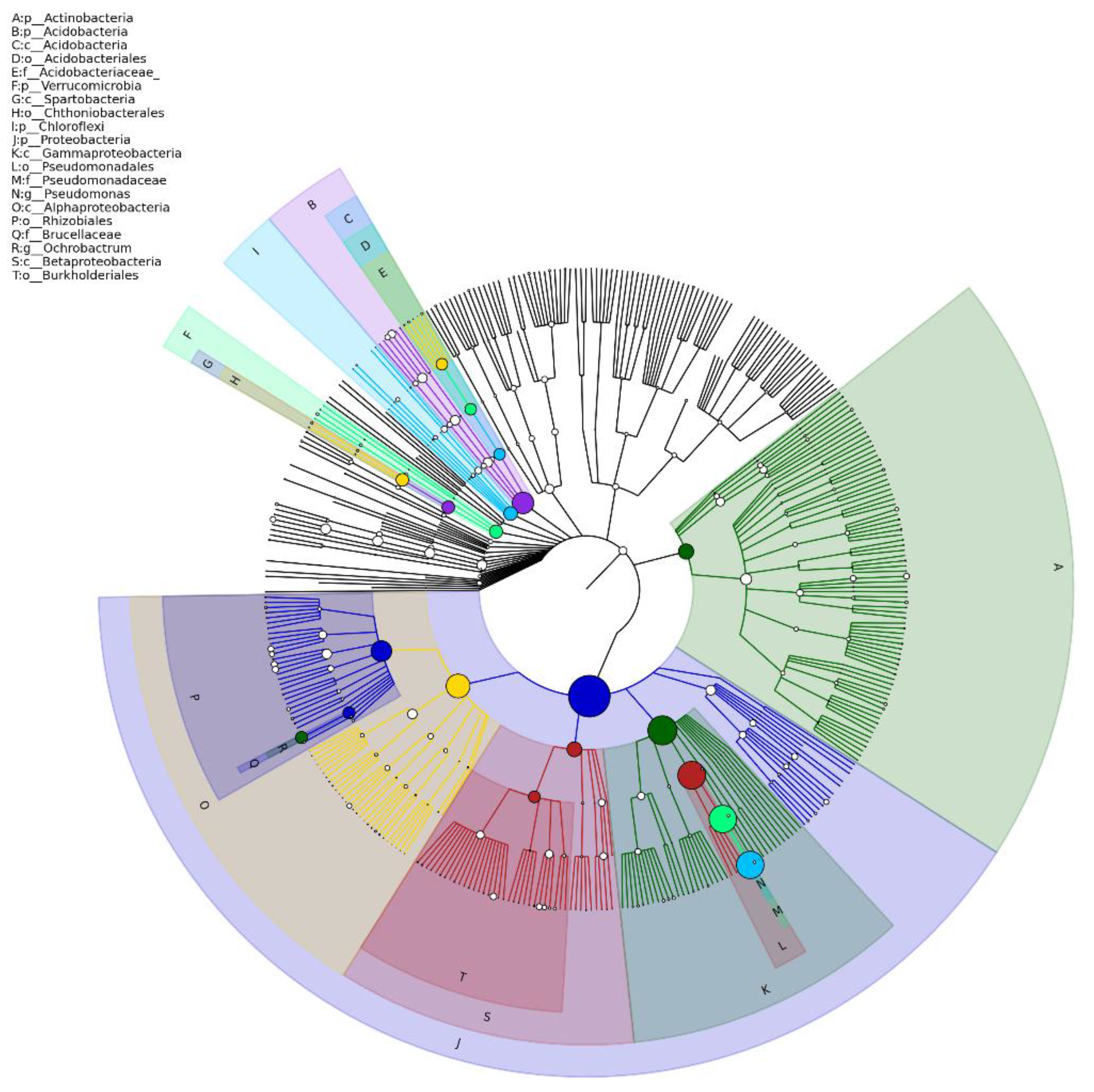
Figure 7.
Taxonomic breakdown of the microbiome of the sampled soil bacterial community based on classified 16S rDNA gene sequences. Representative sequences were taxonomically assigned using the RDP (Ribosomal Database Project) classifier. The tree was generated using GraPhlAn (Graphical Phylogenetic Analysis) [27].
Figure 7.
Taxonomic breakdown of the microbiome of the sampled soil bacterial community based on classified 16S rDNA gene sequences. Representative sequences were taxonomically assigned using the RDP (Ribosomal Database Project) classifier. The tree was generated using GraPhlAn (Graphical Phylogenetic Analysis) [27].

Figure 8.
Venn diagram of common functional groups in the Haplic Cambisols treated with graphene (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1).
Figure 8.
Venn diagram of common functional groups in the Haplic Cambisols treated with graphene (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1).

Figure 9.
Heatmap of bacterial community abundance for KEGG homologous gene clusters (KO), derived from a cluster analysis. The color gradient from green to red indicates increasing similarity. Each cell depicts the similarity for two soil samples in the matrix. (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1; 7, 15, 30, 60, and 90 d mean 7, 15, 30, 60, and 90 days, respectively; K03088: RNA polymerase sigma-70 factor, ECF subfamily; K02014: iron complex outer membrane receptor protein; K03406: methyl-accepting chemotaxis protein; K02004: None; K02035: peptide/nickel transport system substrate-binding protein; K01999: branched-chain amino acid transport system substrate-binding protein; K00059: 3-oxoacyl-[acyl-carrier protein] reductase (EC:1.1.1.100); K00936: None; K01692: enoyl-CoA hydratase (EC:4.2.1.17); K02051: sulfonate/nitrate/taurine transport system substrate-binding protein; K00799: glutathione S-transferase (EC:2.5.1.18); K02050: sulfonate/nitrate/taurine transport system permease protein; K02049: sulfonate/nitrate/taurine transport system ATP-binding protein; K02433: aspartyl-tRNA(Asn)/glutamyl-tRNA (Gln) amidotransferase subunit A (EC:6.3.5.6 6.3.5.7); K07090: None; K00540: None; K03296: hydrophobic/amphiphilic exporter-1 (mainly G- bacteria), HAE1 family; K03704: cold shock protein (beta-ribbon, CspA family); K03559: biopolymer transport protein ExbD; K01996: branched-chain amino acid transport system ATP-binding protein; K01995: branched-chain amino acid transport system ATP-binding protein; K01997: branched-chain amino acid transport system permease protein; K01998: branched-chain amino acid transport system permease protein; K02030: polar amino acid transport system substrate-binding protein; K02029: polar amino acid transport system permease protein; K02057: simple sugar transport system permease protein; K02016: iron complex transport system substrate-binding protein; K03892: ArsR family transcriptional regulator; K01915: glutamine synthetase (EC:6.3.1.2); K00626: acetyl-CoA C-acetyltransferase (EC:2.3.1.9); K00257: None; K02003: None; K06147: ATP-binding cassette, subfamily B, bacterial; K02529: LacI family transcriptional regulator; K09687: antibiotic transport system ATP-binding protein; K01990: ABC-2 type transport system ATP-binding protein; K01784: UDP-glucose 4-epimerase (EC:5.1.3.2); K09686: antibiotic transport system permease protein; K02032: peptide/nickel transport system ATP-binding protein; K02034: peptide/nickel transport system permease protein; K02033: peptide/nickel transport system permease protein; K02027: multiple sugar transport system substrate-binding protein; K02026: multiple sugar transport system permease protein; K02025: multiple sugar transport system permease protein; K08884: serine/threonine protein kinase, bacterial (EC:2.7.11.1); K01834: phosphoglycerate mutase (EC:5.4.2.1); K01897: long-chain acyl-CoA synthetase (EC:6.2.1.3); K02031: peptide/nickel transport system ATP-binding protein; K05366: penicillin-binding protein 1A (EC:2.4.1.3.4.); K01992: ABC-2 type transport system permease protein).
Figure 9.
Heatmap of bacterial community abundance for KEGG homologous gene clusters (KO), derived from a cluster analysis. The color gradient from green to red indicates increasing similarity. Each cell depicts the similarity for two soil samples in the matrix. (A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1; 7, 15, 30, 60, and 90 d mean 7, 15, 30, 60, and 90 days, respectively; K03088: RNA polymerase sigma-70 factor, ECF subfamily; K02014: iron complex outer membrane receptor protein; K03406: methyl-accepting chemotaxis protein; K02004: None; K02035: peptide/nickel transport system substrate-binding protein; K01999: branched-chain amino acid transport system substrate-binding protein; K00059: 3-oxoacyl-[acyl-carrier protein] reductase (EC:1.1.1.100); K00936: None; K01692: enoyl-CoA hydratase (EC:4.2.1.17); K02051: sulfonate/nitrate/taurine transport system substrate-binding protein; K00799: glutathione S-transferase (EC:2.5.1.18); K02050: sulfonate/nitrate/taurine transport system permease protein; K02049: sulfonate/nitrate/taurine transport system ATP-binding protein; K02433: aspartyl-tRNA(Asn)/glutamyl-tRNA (Gln) amidotransferase subunit A (EC:6.3.5.6 6.3.5.7); K07090: None; K00540: None; K03296: hydrophobic/amphiphilic exporter-1 (mainly G- bacteria), HAE1 family; K03704: cold shock protein (beta-ribbon, CspA family); K03559: biopolymer transport protein ExbD; K01996: branched-chain amino acid transport system ATP-binding protein; K01995: branched-chain amino acid transport system ATP-binding protein; K01997: branched-chain amino acid transport system permease protein; K01998: branched-chain amino acid transport system permease protein; K02030: polar amino acid transport system substrate-binding protein; K02029: polar amino acid transport system permease protein; K02057: simple sugar transport system permease protein; K02016: iron complex transport system substrate-binding protein; K03892: ArsR family transcriptional regulator; K01915: glutamine synthetase (EC:6.3.1.2); K00626: acetyl-CoA C-acetyltransferase (EC:2.3.1.9); K00257: None; K02003: None; K06147: ATP-binding cassette, subfamily B, bacterial; K02529: LacI family transcriptional regulator; K09687: antibiotic transport system ATP-binding protein; K01990: ABC-2 type transport system ATP-binding protein; K01784: UDP-glucose 4-epimerase (EC:5.1.3.2); K09686: antibiotic transport system permease protein; K02032: peptide/nickel transport system ATP-binding protein; K02034: peptide/nickel transport system permease protein; K02033: peptide/nickel transport system permease protein; K02027: multiple sugar transport system substrate-binding protein; K02026: multiple sugar transport system permease protein; K02025: multiple sugar transport system permease protein; K08884: serine/threonine protein kinase, bacterial (EC:2.7.11.1); K01834: phosphoglycerate mutase (EC:5.4.2.1); K01897: long-chain acyl-CoA synthetase (EC:6.2.1.3); K02031: peptide/nickel transport system ATP-binding protein; K05366: penicillin-binding protein 1A (EC:2.4.1.3.4.); K01992: ABC-2 type transport system permease protein).

Figure 10.
Ordination plots from a redundancy analysis (RDA) used to explore the relationships between the soil bacterial community and environmental variables. The arrows indicate the lengths and angles between explanatory and response variables and reflect their correlations. Samples from different concentrations of graphene treatments and different incubation times are marked with different colors. (OM = organic matter contents; HN = hydrolytic nitrogen contents; AP = available phosphorus contents; AK = available potassium contents; Dehy = dehydrogenase activities; Cata = catalase activities; Urea = urease activities; Phos = acid phosphatase activities).
Figure 10.
Ordination plots from a redundancy analysis (RDA) used to explore the relationships between the soil bacterial community and environmental variables. The arrows indicate the lengths and angles between explanatory and response variables and reflect their correlations. Samples from different concentrations of graphene treatments and different incubation times are marked with different colors. (OM = organic matter contents; HN = hydrolytic nitrogen contents; AP = available phosphorus contents; AK = available potassium contents; Dehy = dehydrogenase activities; Cata = catalase activities; Urea = urease activities; Phos = acid phosphatase activities).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Selected properties of A1 horizon soil at the initiation of the experiment (mean, n = 3).
| Parameter | pH (KCl) | CEC a (cmol kg−1) b | Organic Matter (%) | Hydrolytic N (mg kg−1) | Available P (mg kg−1) c | Available K (mg kg−1) | Soil Texture |
|---|---|---|---|---|---|---|---|
| Measurement | 4.02 | 40.12 | 4.88 | 282.05 | 61.28 | 304.06 | Loam d |
a Cation exchange capacity; b centimoles of cation charge per kg soil; c extracted with 0.05 mol L−1 HCl–0.025 mol L−1 H2SO4; d according to United States Department of Agriculture Soil Classification.
Table 2.
The experimental layout. Each of the treatments below was determined after 7, 15, 30, 60, or 90 days.
Table 2.
The experimental layout. Each of the treatments below was determined after 7, 15, 30, 60, or 90 days.
| Treatment | A | B | C | D |
|---|---|---|---|---|
| Graphene treatment | - a | + b | + b | + b |
| Graphene concentration (mg·kg−1) | 0 | 10 | 100 | 1000 |
a Without; b with.
Table 3.
Soil bacterial abundance and diversity indices with different concentrations of graphene and different incubation times.
Table 3.
Soil bacterial abundance and diversity indices with different concentrations of graphene and different incubation times.
| Treatment | Chao1 Index | ACE Index | Simpson Index | Shannon Index |
|---|---|---|---|---|
| A 7 days | 4032.65 ± 130.05ab | 4245.73 ±161.62ab | 0.98 ± 0.01a | 10.18 ± 0.04a |
| B 7 days | 3862.37 ± 64.10a | 4050.51 ± 82.35a | 0.97 ± 0.01a | 10.04 ± 0.44a |
| C 7 days | 4243.47 ± 106.09bc | 4460.58 ± 116.62bc | 0.98 ± 0.02a | 10.54 ± 0.86a |
| D 7 days | 4363.79 ± 216.99c | 4560.76 ± 193.13c | 1.00 ± 0.01a | 10.55 ± 0.07a |
| A 15 days | 4200.69 ± 240.42ab | 4478.96 ± 53.54b | 0.99 ± 0.01a | 10.47 ± 0.15ab |
| B 15 days | 3906.09 ± 128.23a | 4079.03 ± 148.81a | 0.99 ± 0.01a | 10.38 ± 0.06ab |
| C 15 days | 4250.09 ± 38.36b | 4383.05 ± 181.02b | 0.99 ± 0.01a | 10.12 ± 0.40a |
| D 15 days | 4368.26 ± 161.44b | 4615.52 ± 55.96b | 1.00 ± 0.00a | 10.63 ± 0.16b |
| A 30 days | 3443.26 ± 198.93ab | 3590.36 ± 75.83ab | 0.99 ± 0.01a | 9.02 ± 0.32a |
| B 30 days | 3781.87 ± 267.92b | 3830.64 ± 99.20b | 0.99 ± 0.00a | 9.33 ± 0 21a |
| C 30 days | 3637.50 ± 20.66b | 3776.57 ± 129.04b | 0.99 ± 0.00a | 9.21 ± 0.61a |
| D 30 days | 3223.14 ± 99.17a | 3385.69 ± 180.84a | 0.99 ± 0.01a | 9.15 ± 0.80a |
| A 60 days | 1531.37 ± 109.87a | 1609.10 ± 128.78a | 0.85 ± 0.01a | 5.75 ± 0.08a |
| B 60 days | 1524.09 ± 38.22a | 1608.36 ± 47.53a | 0.83 ± 0.01a | 5.68 ± 0.12a |
| C 60 days | 1668.98 ± 97.90a | 1791.05 ± 110.48ab | 0.85 ± 0.01a | 5.92 ± 0.06a |
| D 60 days | 1703.83 ± 117.46a | 1830.66 ± 123.96b | 0.86 ± 0.03a | 5.89 ± 0.45a |
| A 90 days | 2829.87 ± 285.22a | 2857.35 ± 44.22a | 0.98 ± 0.01a | 9.95 ± 0.42a |
| B 90 days | 3057.76 ± 47.44a | 3088.18 ± 85.15bc | 1.00 ± 0.01a | 9.67 ± 0.53a |
| C 90 days | 3127.20 ± 192.12a | 3143.73 ±144.95c | 1.00 ± 0.00a | 10.01 ± 0.09a |
| D 90 days | 2894.30 ± 129.88a | 2908.15 ± 85.83ab | 0.99 ± 0.02a | 9.55 ± 0.72a |
A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1. Values followed by the same letter are not statistically different at p < 0.05 by ANOVA (DUNCAN).
Table 4.
Statistical results for Metastats for pairwise comparison test among the different samples (groups).
Table 4.
Statistical results for Metastats for pairwise comparison test among the different samples (groups).
| Group | Phylum | Genus |
|---|---|---|
| A-B | 0 | 38 |
| A-C | 2 | 45 |
| A-D | 1 | 53 |
| B-C | 1 | 32 |
| B-D | 0 | 58 |
| C-D | 1 | 35 |
A: 0 mg kg−1; B: 10 mg kg−1; C: 100 mg kg−1; D: 1000 mg kg−1.
Table 5.
Pearson correlations of organic matter contents (OM), pH values (pH), hydrolytic nitrogen contents (HN), available phosphorus contents (AP), available potassium contents (AK), dehydrogenase activities (Dehy), catalase activities (Cata), urease activities (Urea), and acid phosphatase activities (Phos).
Table 5.
Pearson correlations of organic matter contents (OM), pH values (pH), hydrolytic nitrogen contents (HN), available phosphorus contents (AP), available potassium contents (AK), dehydrogenase activities (Dehy), catalase activities (Cata), urease activities (Urea), and acid phosphatase activities (Phos).
| OM | pH | HN | AP | AK | Dehy | Cata | Urea | |
|---|---|---|---|---|---|---|---|---|
| pH | 0.157 | |||||||
| HN | 0.152 | 0.649 ** | ||||||
| AP | −0.255 * | −0.561 ** | −0.295 ** | |||||
| AK | 0.014 | 0.598 ** | 0.698 ** | −0.067 | ||||
| Dehy | 0.164 | 0.320 * | 0.172 | −0.171 | 0.258 * | |||
| Cata | 0.167 ** | −0.548 ** | −0.660 ** | 0.305 * | −0.592 ** | −0.183 | ||
| Urea | 0.191 | 0.352 ** | 0.708 ** | −0.335 ** | 0.527 ** | 0.094 | −0.622 ** | |
| Phos | 0.004 | 0.216 | 0.298 * | −0.034 ** | 0.198 | −0.023 | −0.128 | 0.140 |
* p < 0.05; ** p < 0.01.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Song, J.; Duan, C.; Sang, Y.; Wu, S.; Ru, J.; Cui, X. Effects of Graphene on Bacterial Community Diversity and Soil Environments of Haplic Cambisols in Northeast China. Forests 2018, 9, 677. https://doi.org/10.3390/f9110677
AMA Style
Song J, Duan C, Sang Y, Wu S, Ru J, Cui X. Effects of Graphene on Bacterial Community Diversity and Soil Environments of Haplic Cambisols in Northeast China. Forests. 2018; 9(11):677. https://doi.org/10.3390/f9110677
Chicago/Turabian StyleSong, Jinfeng, Chengwei Duan, Ying Sang, Shaoping Wu, Jiaxin Ru, and Xiaoyang Cui. 2018. "Effects of Graphene on Bacterial Community Diversity and Soil Environments of Haplic Cambisols in Northeast China" Forests 9, no. 11: 677. https://doi.org/10.3390/f9110677
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.