Chemical Diversity and Defence Metabolism: How Plants Cope with Pathogens and Ozone Pollution


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Plant Secondary Metabolism
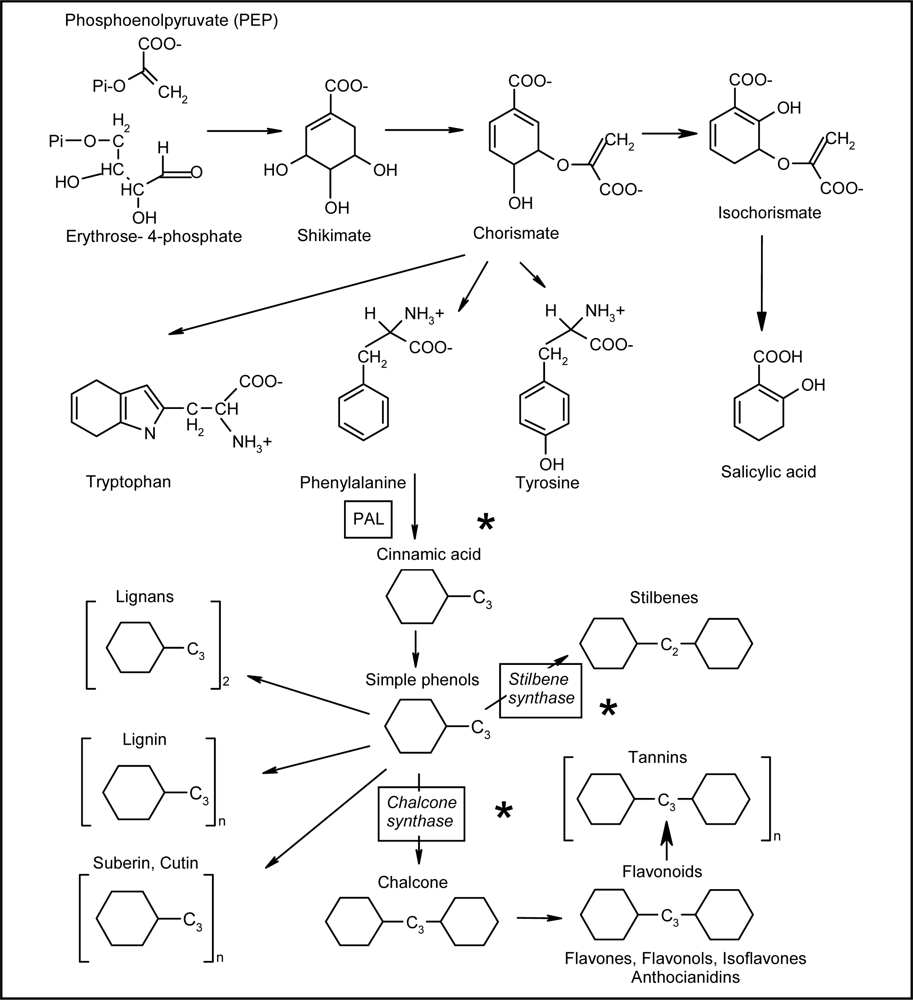
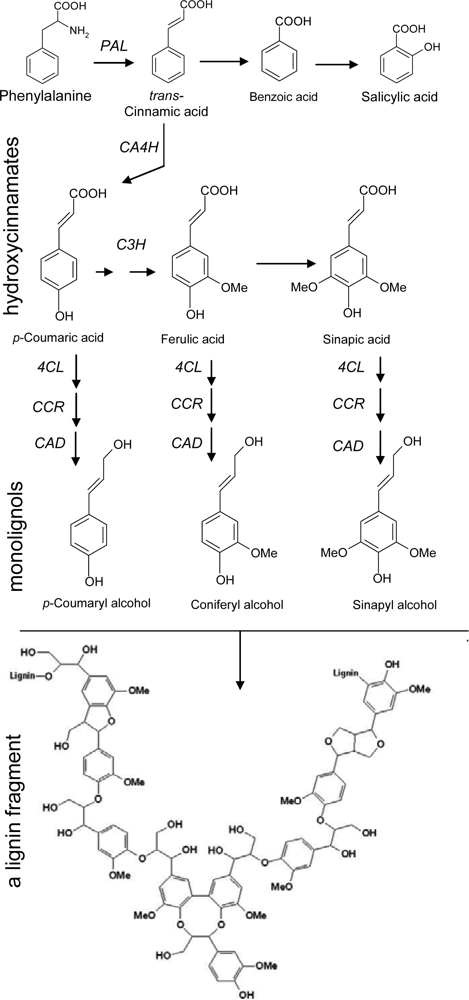
2.1. Phenylpropanoid Pathway
2.2. Isoprenoid Pathway
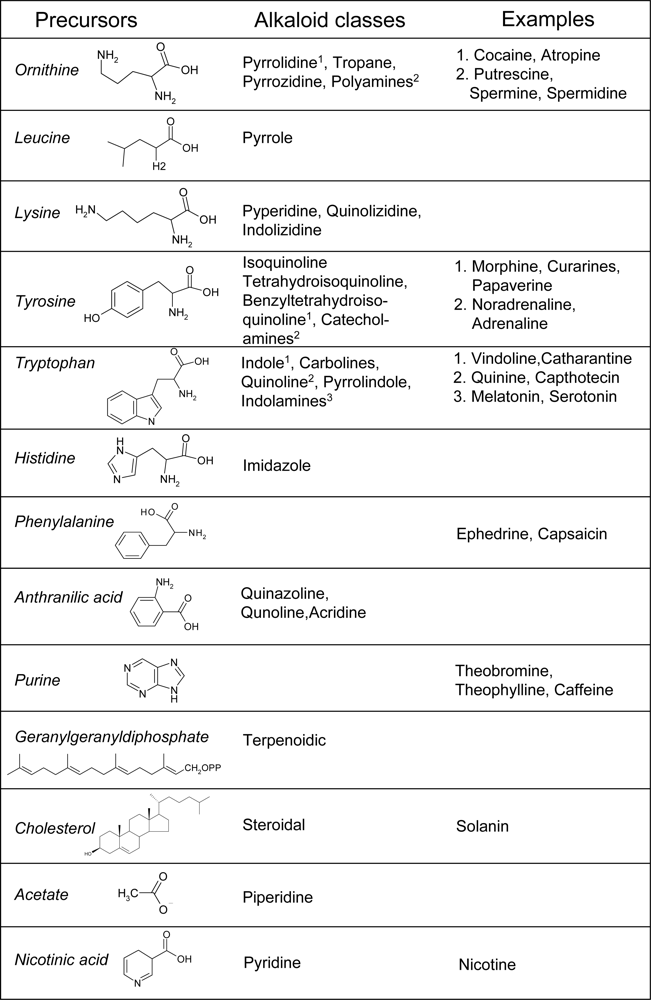
2.3. Alkaloid Pathways
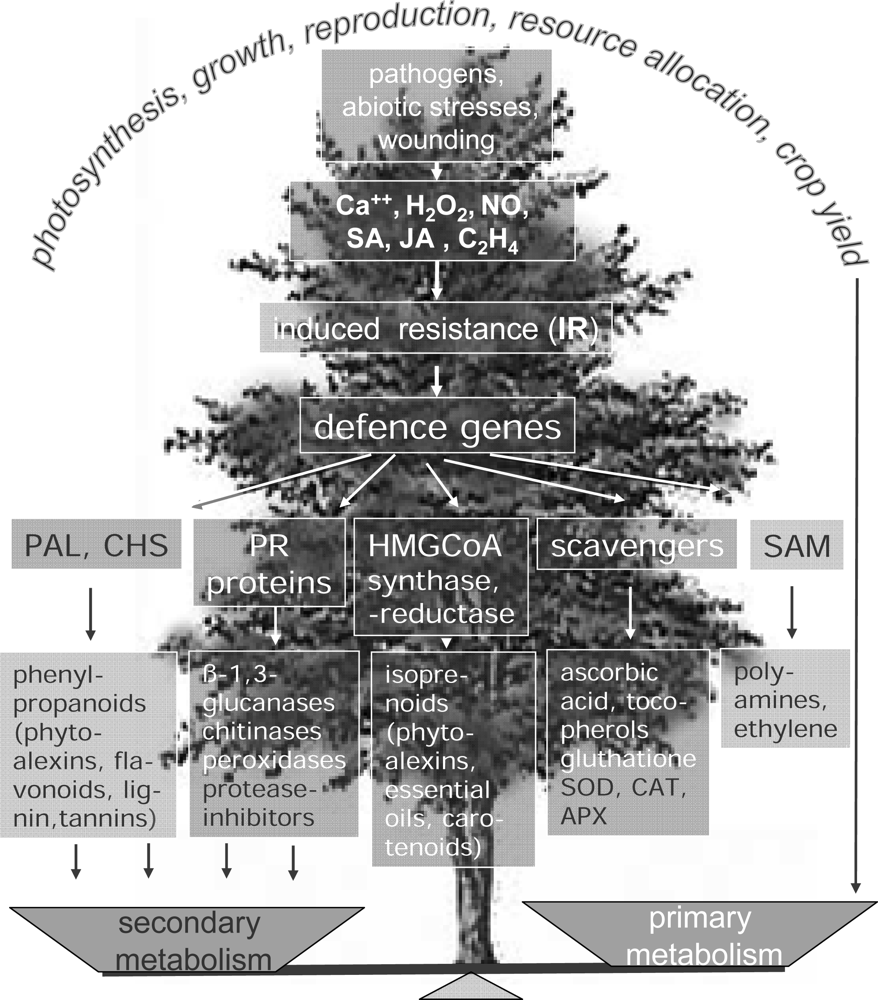
3. Secondary Metabolites Involved in Plant Resistance against Pathogens
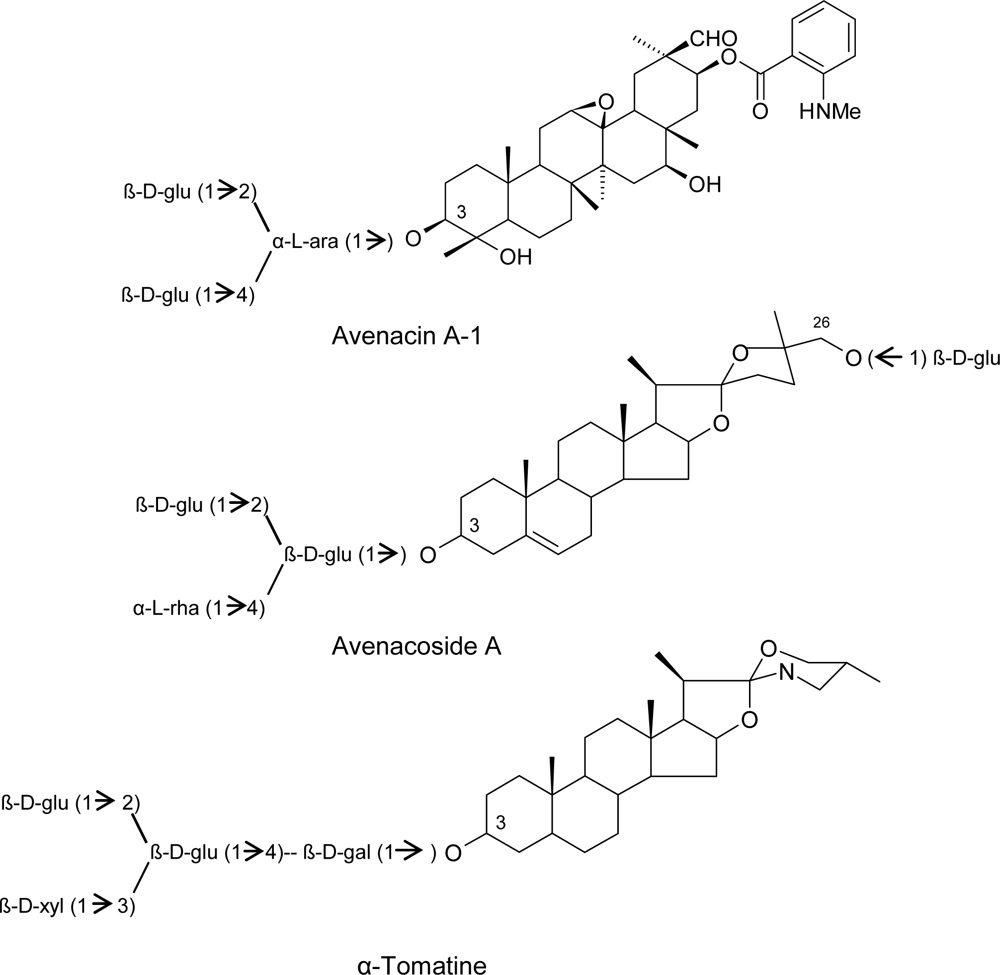
3.1. Phytoanticipins
3.2. Phytoalexins
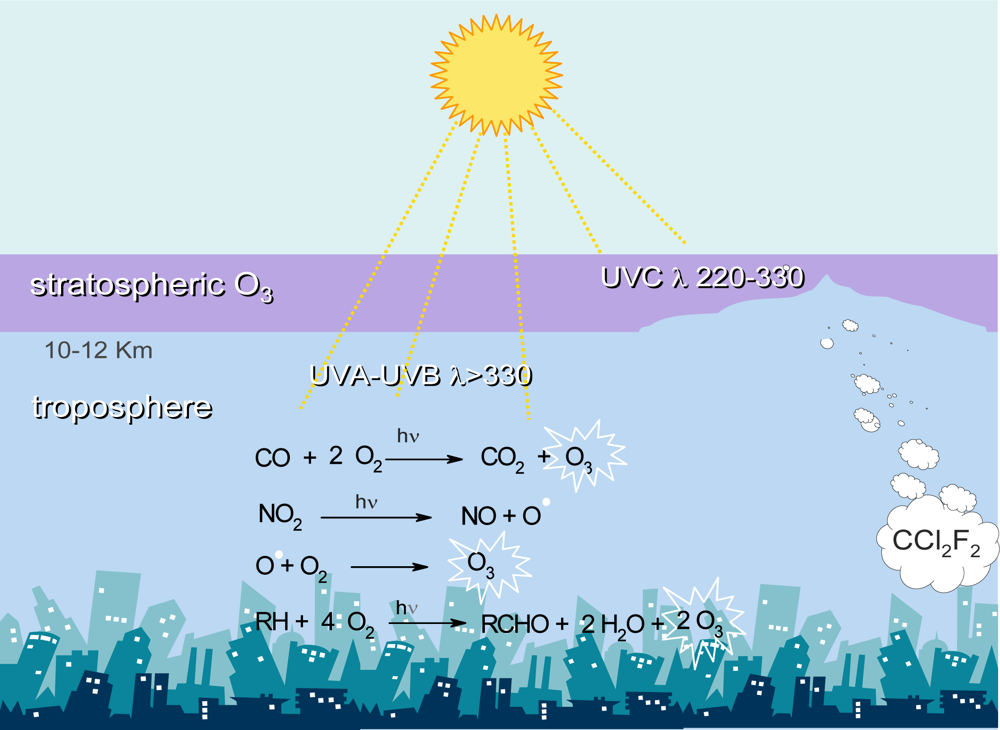
4. Tropospheric Ozone and Plant Health
4.1. Ozone and Phenylpropanoids
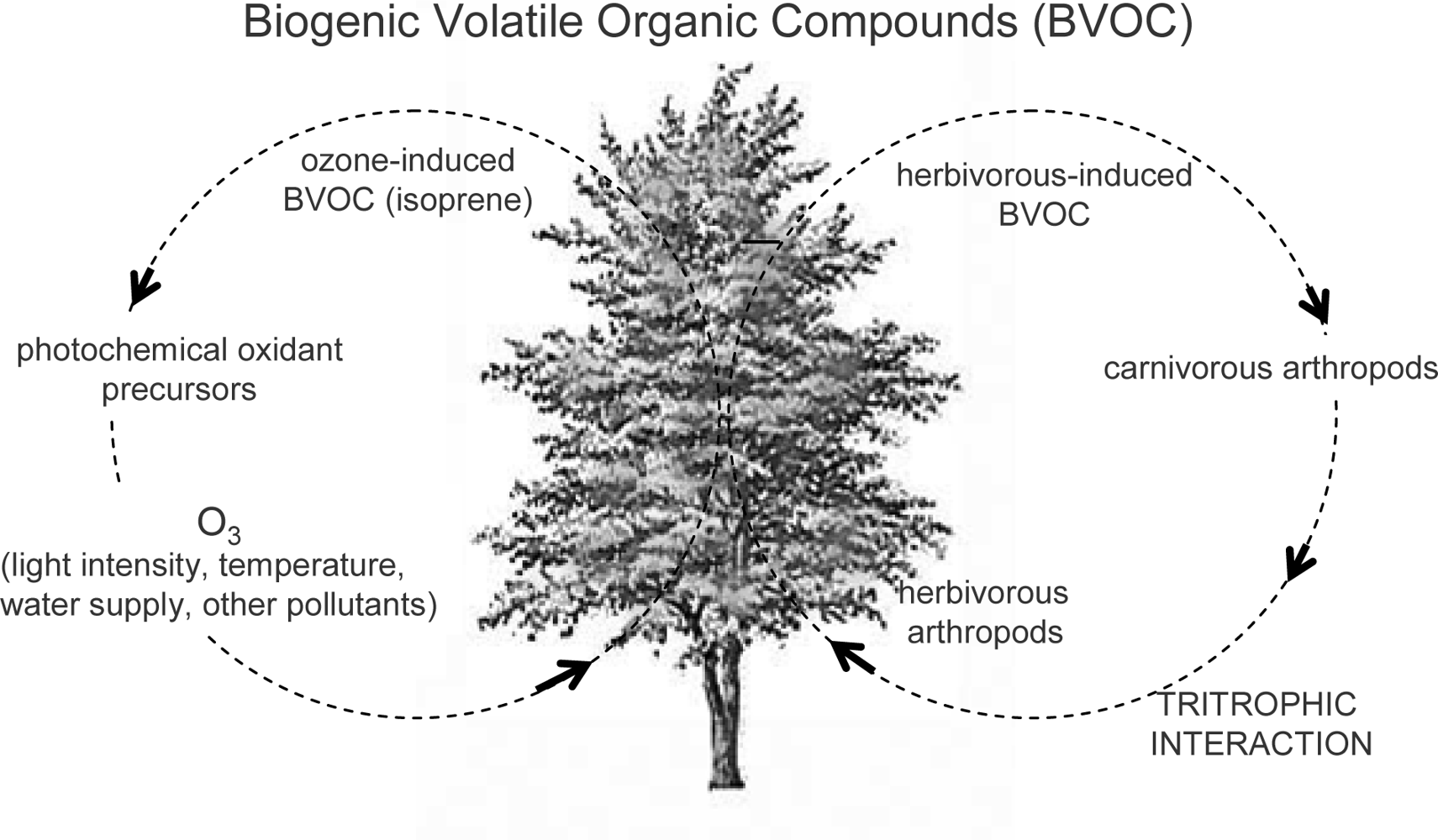
4.2. Ozone and Isoprenoids
4.3. Ozone and Alkaloids
5. Conclusions
References and Notes
- Lichtenthaler, HK. The stress concept in plants: An introduction. Ann. N.Y. Acad. Sci 1998, 851, 187–198. [Google Scholar]
- Heiser, I; Elstner, EF. The biochemistry of plant stress and disease. Ann. N.Y. Acad. Sci 1998, 851, 224–232. [Google Scholar]
- Firn, RD; Jones, CG. A Darwinian view of metabolism: Molecular properties determine fitness. J. Exp. Bot 2009, 60, 719–726. [Google Scholar]
- Paiva, N. An introduction to the biosynthesis of chemicals used in plant-microbe communication. J. Plant Growth Regul 2000, 19, 131–143. [Google Scholar]
- Facchini, PJ. Alkaloid biosynthesis in plants: Biochemistry, cell biology, molecular regulation and metabolic engineering. Annu. Rev. Plant Physiol. Plant Mol. Biol 2001, 52, 29–66. [Google Scholar]
- Holstein, SA; Hohl, RJ. Isoprenoids: Remarkable diversity of form and function. Lipids 2004, 34, 293–309. [Google Scholar]
- Iriti, M; Faoro, F. Plant defense and human nutrition: the phenylpropanoids on the menu. Curr. Top. Nutrac. Res 2004, 2, 47–95. [Google Scholar]
- Weaver, LM; Hermann, KM. Dynamics of the shikimate pathway in plants. Tends Plant Sci 1997, 2, 346–351. [Google Scholar]
- Hahlbrock, K; Scheel, D. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol 1989, 40, 347–369. [Google Scholar]
- Wildermuth, MC; Dewdney, J; Wu, G; Ausubel, FM. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 2001, 414, 562–565. [Google Scholar]
- Iriti, M; Faoro, F. Lipid biosynthesis in Spermatophyta. In Floriculture, Ornamental and Plant Biotechnology; Teixeira da Silva, JA, Ed.; Global Science Books: London, UK, 2006; Volume 1, Chapter 40; p. 359. [Google Scholar]
- Sacchettini, JC; Poulter, CD. Creating isoprenoid diversity. Science 1997, 277, 1788–1789. [Google Scholar]
- Chappell, J. The biochemistry and molecular biology of isoprenoid metabolism. Plant Physiol 1995, 107, 1–6. [Google Scholar]
- Bach, TJ. Synthesis and metabolism of mevalonic acid in plants. Plant Physiol. Biochem 1987, 25, 163–178. [Google Scholar]
- Poulter, CD; Rilling, HC. The prenyl transferase reaction, enzymatic and mechanistic studies of the 1′-4 coupling reaction in the terpene biosynthetic pathway. Acc. Chem. Res 1978, 1, 307–313. [Google Scholar]
- Croteau, R. Biosynthesis and catabolism of monoterpenoids. Chem. Rev 1987, 87, 929–954. [Google Scholar]
- Cane, DE. Enzymatic formation of sesquiterpenes. Chem. Rev 1990, 90, 1089–1103. [Google Scholar]
- Dogbo, A; Camara, B. Purification of isopentenyl pyrophosphate isomerase and geranylgeranyl pyrophosphate synthase from Caspicum chromoplasts by affinity chromatography. Biochim. Biophys. Acta 1987, 92, 140–148. [Google Scholar]
- Abe, I; Rohmer, M; Prestwich, GD. Enzymatic cyclization of squalene and oxidosqualene to sterols and triterpens. Chem. Rev 1993, 93, 2189–2206. [Google Scholar]
- Wendt, KU; Schulz, GE. Isoprenoid biosynthesis: manifold chemistry catalysed by similar enzymes. Curr. Biol 1998, 6, 127–133. [Google Scholar]
- Cordell, GA; Quinn-Beattie, ML; Farsnsworth, NR. The potential of alkaloids in drug discovery. Phytot. Res 2001, 15, 183–205. [Google Scholar]
- Hughes, EH; Shanks, JV. Metabolic engineering of plants for alkaloid production. Metab. Engin 2002, 4, 41–48. [Google Scholar]
- Iriti, M; Faoro, F. Review on innate and specific immunity in plants and animals. Mycopathologia 2007, 164, 57–64. [Google Scholar]
- VanEtten, HD; Mansfield, JW; Bailey, JA; Farmer, EE. Two classes of plant antibiotics: Phytoalexins versus ‘phytoanticipins’. Plant Cell 1994, 6, 1191–1192. [Google Scholar]
- Osbourn, AE. Preformed antimicrobial compounds and plant defence against fungal attack. Plant Cell 1996, 8, 1821–1831. [Google Scholar]
- Price, KR; Johnson, IT; Fenwick, GR. The chemistry and biological significance of saponins in food and feedingstuffs. Crit. Rev. Food Sci. Nutr 1987, 26, 27–133. [Google Scholar]
- Osbourn, AE. Saponins and plant defence-A soap story. Trends Plant Sci 1996, 1, 4–9. [Google Scholar]
- Mansfield, JW. Antimicrobial Compounds. In Mechanisms of Resistance to Plant Diseases; Slusarenko, AJ, Fraser, RSS, van Loon, LC, Eds.; Kluwer Academic Press: Dordrecht, The Netherlands, 2000. [Google Scholar]
- Turner, EM. The nature of resistance of oats to the take-all fungus. J. Exp. Bot 1953, 4, 264–271. [Google Scholar]
- Crombie, L; Crombie, WML; Whiting, DA. Structures of the four avenacins, oat root resistance factors to take-all disease. J. Chem. Soc., Chem. Commun 1984, 4, 246–248. [Google Scholar]
- Crombie, WML; Crombie, L. Distribution of avenacins A-1, A-2, B-1 and B-2 in oat roots: Their fungicidal activity toward ‘take-all’ fungus. Phytochemistry 1986, 25, 2069–2073. [Google Scholar]
- Osbourn, AE; Clarke, BR; Lunness, P; Scott, PR; Daniels, MJ. An oat species lacking avenacin is susceptible to infection by Gaeumannomyces graminis var. tritici. Physiol. Mol. Plant Pathol 1994, 45, 457–467. [Google Scholar]
- Osbourn, AE; Clarke, BR; Dow, JM; Daniels, MJ. Partial characterization of avenacinase from Gaeumannomyces graminis var.avenae. Physiol. Mol. Plant Pathol 1991, 38, 301–312. [Google Scholar]
- Osbourn, AE. Preformed antimicrobial compounds and plant defense against fungal attack. Plant Cell 1996, 8, 1821–1831. [Google Scholar]
- Dow, JM; Callow, JA. A possible role for α-tomatine in the varietal-specific resistance of tomato to Cladosporium fulvum. Phytopathol. Z 1978, 92, 211–216. [Google Scholar]
- Smith, CA; MacHardy, WE. The significance of tomatine in the host response of susceptible and resistant tomato isolines infected with two races of Fusarium oxysporum f. sp. lycopersici. Phytopathology 1982, 72, 415–419. [Google Scholar]
- Pegg, GF; Woodward, S. Synthesis and metabolism of α-tomatine in tomato isolines in relation to resistance to Verticillium albo-atrum. Physiol. Mol. Plant Pathol 1986, 28, 187–201. [Google Scholar]
- Zagrobelny, M; Bak, S; Møller, BM. Cyanogenesis in plants and arthropods. Phytochemistry 2008, 69, 1457–1468. [Google Scholar]
- Fenwick, GR; Heaney, RK; Mullin, WJ. Glucosinolates and their breakdown products in food and food plants. Crit. Rev. Food Sci. Nutr 1983, 18, 123–301. [Google Scholar]
- Nahrstedt, A. Cyanogenic compounds as protecting agents for organisms. Plant Syst. Evol 1985, 150, 35–47. [Google Scholar]
- Poulton, JE; Li, CP. Tissue level compartmentation of (R)-amygdalin and amygdalin hydrolase prevents large-scale cyanogenesis in undamaged Prunus seeds. Plant Physiol 1994, 104, 29–35. [Google Scholar]
- Swain, E; Poulton, JE. Utilization of amygdalin during seedling development of Prunus serotina. Plant Physiol 1994, 106, 437–445. [Google Scholar]
- Wang, P; Matthews, DE; VanEtten, HD. Purification and characterization of cyanide hydratase from the phytopathogenic fungus Gloeocercospora sorghi. Arch. Biochem. Biophys 1992, 298, 569–575. [Google Scholar]
- Benderoth, M; Textor, S; Windsor, AJ; Mitchell-Olds, T; Gershenzon, J; Kroymann, J. Positive selection driving diversification in plant secondary metabolism. Proc. Nat. Acad. Sci. USA 2006, 109, 9118–9123. [Google Scholar]
- Bidart-Bouzat, MG; Kliebenstein, DJ. Differential levels of insect herbivory in the field associated with genotypic variation in glucosinolates in Arabidopsis thaliana. J. Chem. Ecol 2008, 34, 1026–1037. [Google Scholar]
- Mithen, R. Leaf glucosinolate profile and their relationship to pest and disease resistance in oilseed rape. Euphytica 1992, 63, 71–83. [Google Scholar]
- Lenman, M; Falk, A; Xue, J; Rask, L. Characterization of a Brassica napus myrosinase pseudogene: myrosinases are members of the BGA family of β-glycosidases. Plant Mol. Biol 1993, 21, 463–474. [Google Scholar]
- Greenhalgh, JG; Mitchell, ND. The involvement of flavour volatiles in the resistance to downy mildew of wild and cultivated forms of Brassica oleracea. New Phytol 1976, 77, 391–398. [Google Scholar]
- Mithen, R; Magrath, R. Glucosinolates and resistance to Leptosphaeria maculans in wild and cultivated Brassiaca species. Plant Breed 1992, 108, 60–68. [Google Scholar]
- Harthill, WFT; Sutton, PG. Inhibition of Mycosphaerella brassicola ascospores on young cabbage and cauliflower leaves. Ann. Appl. Biol 1980, 96, 153–161. [Google Scholar]
- Mert-Türk, F. Phytoalexins: Defence or just a response to stress? J. Cell Mol. Biol 2002, 1, 1–6. [Google Scholar]
- Grayer, RJ; Harborne, JJ. A survey of antifungal compounds from higher plants. Phytochemistry 1994, 37, 19–42. [Google Scholar]
- Dixon, RA; Paiva, NL. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar]
- Bailey, JA; Rowell, PM; Arnold, GM. The temporal relationship between infected cell death, phytoalexin accumulation and the inhibition of hyphal development during resistance of Phaseolus vulgaris to Colletotrichum lindemuthianum. Physiol. Plant Pathol 1980, 1, 435–449. [Google Scholar]
- Blount, JW; Dixon, RA; Paiva, NL. Stress responses in alfalfa (Medicago sativa L.) XVI. Antifungal activity of medicarpin and its biosynthetic precursors: Implications for the genetic manipulation of stress metabolites. Physiol. Mol. Plant Pathol 1993, 41, 333–349. [Google Scholar]
- DiCenzo, GL; VanEtten, HD. Studies on the late steps of (+) pisatin biosynthesis: Evidence for (−) enantiomeric intermediates. Phytochemistry 2006, 67, 675–683. [Google Scholar]
- Jeandet, P; Douillet-Breuil, AC; Bessis, R; Debord, S; Sbaghi, M; Adrian, M. Phytoalexins from the Vitaceae: Biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity and metabolism. J. Agric. Food Chem 2002, 50, 2731–2741. [Google Scholar]
- Jeandet, P; Sbaghi, M; Bessis, R; Meunier, P. The potential relationship of stilbene (resveratrol) synthesis to anthocyanin content in grape berry skins. Vitis 1995, 34, 91–94. [Google Scholar]
- Iriti, M; Rossoni, M; Borgo, M; Faoro, F. Benzothiadiazole enhances resveratrol and anthocyanin biosynthesis in grapevine, meanwhile improving resistance to Botrytis cinerea. J. Agric. Food Chem 2004, 52, 4406–4413. [Google Scholar]
- Glawischnig, E; Hansen, BG; Olsen, CE; Halkier, BA. Camalexin is synthesized from indole-3-acetaldoxime, a key branching point between primary and secondary metabolism in Arabidopsis. Proc. Nat. Acad. Sci. USA 2004, 101, 8245–8250. [Google Scholar]
- Dutsch, HV. Vertical ozone distribution on a global scale. Pure Appl. Geophys 1978, 116, 511–529. [Google Scholar]
- Kerr, JB; McElroy, CT. Evidence for large trend of ultraviolet-B radiation linked to ozone depletion. Science 1993, 262, 1032–1034. [Google Scholar]
- Logan, JA. Tropospheric ozone: seasonal behaviour, trends and anthropogenic influences. J. Geophys. Res 1985, 90, 10463–10482. [Google Scholar]
- Crutzen, PJ; Lelieveld, J. Human impacts on atmospheric chemistry. Annu. Rev. Earth Planet. Sci 2001, 29, 17–45. [Google Scholar]
- Fowler, D; Cape, JN; Coyle, M; Smith, RI; Hjellbrekke, A-G; Simpson, D; Derwent, RG; Johnson, CE. Modelling photochemical oxidant formation, transport, deposition and exposure of terrestrial ecosystems. Environ. Pollut 1999, 100, 43–55. [Google Scholar]
- Kley, D; Kleinmann, M; Sandermann, H; Krupa, S. Photochemical oxidants: State of the science. Environ. Pollut 1999, 100, 19–42. [Google Scholar]
- Mustafa, MG. Biochemical basis of ozone toxicity. Free Radical Biol. Med 1990, 9, 245–265. [Google Scholar]
- Kelly, FJ; Mudway, I; Krishna, MT; Holgate, ST. The free radical basis of air pollution: Focus on ozone. Resp. Med 1995, 89, 647–656. [Google Scholar]
- Pryor, WA; Das, B; Church, DF. The ozonation of unsaturated fatty acids: Aldehydes and hydrogen peroxide as products and possible mediators of ozone toxicity. Chem. Res. Toxicol 1991, 4, 341–348. [Google Scholar]
- Mudd, JB; Leavitt, R; Ongun, A; McManus, TT. Reaction of ozone with amino acids and proteins. Atmos. Environ 1969, 3, 669–682. [Google Scholar]
- Hussain, MZ; Mustafa, MG; Ghani, QP; Bhatnagar, RS. Stimulation of poly(ADP-ribose) synthetase activity in the lungs of mice exposed to a low level of ozone. Archiv. Biochem. Biophys 1985, 241, 477–485. [Google Scholar]
- Adams-Phillips, L; Wan, J; Tan, X; Dunning, FM; Meyers, BC; Michelmore, RW; Bent, AF. Discovery of ADP-Ribosylation and other plant defense pathway elements through expression profiling of four different Arabidopsis–Pseudomonas R-avr interactions. Mol. Plant-Microbe Interact 2008, 21, 646–657. [Google Scholar]
- Bergmann, E; Bender, J; Weigel, J. Ozone threshold doses and exposure-response relationships for the development of ozone injury symptoms in wild plant species. New Phytol 1999, 144, 423–435. [Google Scholar]
- Krupa, S; McGrath, MT; Andersen, CP; Booker, F; Burkey, KO; Chappelka, AH; Chevone, BI; Pell, EJ; Zilinskas, BA. Ambient ozone and plant health. Plant Dis 2000, 85, 4–12. [Google Scholar]
- Torsethaugen, G; Pell, EJ; Assmann, SM. Ozone inhibits guard cell K+ channels implicated in stomatal opening. Proc. Nat. Acad. Sci. USA 1999, 96, 13577–13582. [Google Scholar]
- Black, VJ; Black, CR; Roberts, JA; Stewart, CA. Impact of ozone on the reproductive development of plants. New Phytol 2000, 147, 421–447. [Google Scholar]
- Enyedi, AJ; Eckardt, NA; Pell, EJ. Activity of ribulose bisphosphate carboxylase/oxygenase from potato cultivars with differential response to ozone stress. New Phytol 1992, 122, 493–500. [Google Scholar]
- Reddy, GN; Arteca, RN; Dai, YR; Flores, HE; Negram, FB; Pell, EJ. Changes in ethylene and polyamines in relation to mRNA levels of the large and small subunits of ribulose bisphosphate oxygenase in ozone stressed potato foliage. Plant Cell Environ 1993, 120, 819–826. [Google Scholar]
- Guidi, L; Degl’Innocenti, E; Martinelli, F; Piras, M. Ozone effects on carbon metabolism in sensitive and insensitive Phaseolus vulgaris cultivars. Environ. Exp. Bot 2009, 66, 117–125. [Google Scholar]
- Toumainen, J; Pellinen, R; Roy, S; Kiiskinen, M; Eloranta, T; Karjalainen, R; Kangasjärvi, J. Ozone affect birch (Betula pendula Roth) phenylpropanoid, polyamine and reactive oxygen detoxifying pathways at biochemical and gene expression levels. J. Plant Physiol 1996, 148, 179–188. [Google Scholar]
- Keen, NT; Taylor, OC. Ozone injury in soybeans: Isoflavonoid accumulation is related to necrosis. Plant Physiol 1975, 55, 731–733. [Google Scholar]
- Sharma, YK; Davis, KR. Ozone-induced expression of stress-related genes in Arabidopsis thaliana. Plant Physiol 1994, 105, 1089–1096. [Google Scholar]
- Eckey-Kaltenbach, HE; Ernst, D; Heller, W; Sandermann, H. Biochemical plant response to ozone IV. Cross-induction of defensive pathways in parsley (Petroselinum crispum L.) plants. Plant Physiol 1994, 104, 67–74. [Google Scholar]
- Dercks, W; Trumble, JT; Winter, C. Impact of atmospheric pollution on linear furanocoumarin in celery. J. Chem. Ecol 1990, 16, 443–454. [Google Scholar]
- Rao, MV; Davis, KR. The physiology of ozone-induced cell death. Planta 2001, 213, 682–690. [Google Scholar]
- Heiden, AC; Hoffmann, T; Kahl, J; Kley, D; Klockow, D; Langebartels, C; Mehlhorn, H; Sandermann, H; Schraudner, M; Schuh, G; Wildt, J. Emission of volatile organic compounds from ozone-exposed plants. Ecolog. Appl 1999, 9, 1160–1167. [Google Scholar]
- Sharma, YK; Leon, J; Raskin, I; Davis, KR. Ozone-induced responses in Arabidopsis thaliana: The role of salicylic acid in the accumulation of defense-related transcripts and induced resistance. Proc. Nat. Acad. Sci. USA 1996, 93, 5099–5104. [Google Scholar]
- Langebartels, C; Kerner, K; Leonardi, S; Schraudner, M; Trost, M; Heller, W; Sandermann, H. Biochemical plant resonses to ozone I: Differential induction of polyamine and ethylene biosynthesis in tobacco. Plant Physiol 1991, 9, 882–889. [Google Scholar]
- Faoro, F; Iriti, M. Plant cell death and cellular alterations induced by ozone: Key studies in Mediterranean conditions. Environ. Pollut 2009, 157, 1470–1477. [Google Scholar]
- Schraudner, M; Ernst, D; Langebartels, C; Sandermann, H. Biochemical plant responses to ozone III. Activation of the defence-related proteins β-1,3-glucanase and chitinase in tobacco leaves. Plant Physiol 1992, 99, 1321–1328. [Google Scholar]
- Thalmair, M; Bauw, G; Thiel, S; Döhring, T; Langebartels, C; Sandermann, H. Ozone and ultraviolet B effects on the defense-related proteins β-1,3-glucanase and chitinase in tobacco. J. Plant Physiol 1996, 148, 222–228. [Google Scholar]
- Pääkkönen, E; Seppänen, S; Holpainen, T; Kokko, H; Kärenlampi, S; Kärenlampi, L; Kangasjärvi, L. Induction of genes for stress proteins PR-10 and PAL in relation to growth, visible injury and stomatal conductance in birch (Betula pendula) clones exposed to ozone and/or drought. New Phytol 1998, 138, 295–305. [Google Scholar]
- Paolacci, AR; D’Ovidio, R; Marabottini, R; Nali, C; Lorenzini, G; Abenavoli, MR; Badiani, M. Ozone induces a differential accumulation of phenylalanine ammonia-lyase, chalcone synthase and chalcone isomerase RNA transcripts in sensitive and resistant bean cultivars. Aust. J. Plant Physiol 2001, 28, 425–428. [Google Scholar]
- Schubert, R; Fischer, R; Hain, R; Schreier, PH; Bahnweg, D; Ernst, H; Sandermann, H. An ozone-responsive region of the grapevine resveratrol synthase promoter differs from the basal pathogen-responsive sequence. Plant Mol. Biol 1997, 34, 417–436. [Google Scholar]
- Sgarbi, E; Baroni Fornasiero, R; Lins, AP; Medeghini Bonatti, P. Phenol metabolism in differentially affected by ozone in two cell lines from grape (Vitis vinifera L.) leaf. Plant Sci 2003, 165, 951–957. [Google Scholar]
- Foy, D; Lee, EH; Rowland, R; Devine, TE; Buzzel, RI. Ozone tolerance related to flavonol glycoside genes in soybean. J. Plant Nutr 1995, 18, 637–647. [Google Scholar]
- Saleem, A; Loponen, Y; Pihlaja, K; Oksanen, E. Effects of long-term open-field ozone exposure on leaf phenolics of european silver birch (Betula pendula Roth). J. Chem. Ecol 2001, 27, 1049–1062. [Google Scholar]
- Karonen, M; Ossipov, V; Ossopova, S; Kapari, L; Loponen, J; Matsumura, H; Kohno, T; Mikami, C; Sakai, Y; Izuta, T; Pihlaja, K. Effects of elevated carbon dioxide and ozone on foliar proanthocyanidins in Betula platyphylla, Betula ermanii and Fagus crenata seedlings. J. Chem. Ecol 2006, 32, 1445–1458. [Google Scholar]
- Cabané, M; Pireaux, J-C; Léger, E; Weber, E; Dizengremel, P; Pollet, B; Lapierre, C. Condensed lignins are synthesized in poplar leaves exposed to ozone. Plant Physiol 2004, 134, 586–594. [Google Scholar]
- Guidi, L; Degl’Innocenti, E; Genovesi, S; Soldatini, GF. Photosynthetic process and activities of enzymes involved in the phenylpropanoid pathway in resistant and sensitive genotypes of Lycopersicon esculentum L. exposed to ozone. Plant Sci 2005, 168, 153–160. [Google Scholar]
- Galliano, H; Heller, W; Sandermann, H. Ozone induction and purification of spruce cinnamyl alcohol dehydrogenase. Phytochemistry 1993, 32, 557–563. [Google Scholar]
- Tomlinson, H; Rich, S. Effect of ozone on sterols and sterol derivatives in bean leaves. Phytopathology 1971, 61, 1404–1405. [Google Scholar]
- Trevathan, LE; Moore, LD; Orcutt, DM. Symptom expression and free sterol and fatty acid composition of flue-cured tobacco plants exposed to ozone. Phytopathology 1979, 69, 582–585. [Google Scholar]
- Grunwald, C; Endress, AG. Foliar sterols in soybeans exposed to chronic levels of ozone. Plant Physiol 1985, 77, 245–247. [Google Scholar]
- Grunwald, C. Effects of free sterols, steryl ester and steryl glycoside in membrane permeability. Plant Physiol 1971, 48, 653–655. [Google Scholar]
- Evans, LS; Ting, IP. Ozone-induced membrane permeability changes. Am. J. Bot 1973, 60, 155–162. [Google Scholar]
- Grunwald, C. Effects of free sterols, steryl ester and steryl glycoside in membrane permeability. Plant Physiol 1974, 54, 624–628. [Google Scholar]
- Menser, HA; Chaplin, JF; Cheng, ALS; Sorokin, T. Polyphenols, phytosterols and reduced sugar in air-cured tobacco leaves injured by ozone air pollution. Tobacco Sci 1977, 21, 35–38. [Google Scholar]
- Wegener, A; Gimbel, W; Werner, T; Hani, J; Ernst, D; Sandermann, H. Molecular cloning of ozone-inducible protein from Pinus sylvestris L. with high sequence similarity to vertebrate 3-hydroxy-3-methylglutaryl-CoA-synthase. Biochim. Biophys. Acta 1997, 1350, 247–252. [Google Scholar]
- Cheng, A-X; Lou, Y-G; Mao, Y-B; Lu, S; Wang, L-J; Chen, X-Y. Plant terpenoids: Biosynthesis and ecological functions. J. Integr. Plant Biol 2007, 49, 179–186. [Google Scholar]
- Shamay, Y; Raskin, VI; Brandis, AS; Steinberger, HE; Marder, JB; Schwartz, A. Ozone treatment affects pigment precursor metabolism in pine seedlings. Physiol. Plant 2001, 112, 285–292. [Google Scholar]
- Kesselmeier, J; Staudt, M. Biogenic volatile organic compounds (VOC): An overview on emission, physiology and ecology. J. Atmos. Chem 1999, 33, 23–88. [Google Scholar]
- Dicke, M. Chemical ecology of host-plant selection by herbivorous arthropods: A multitrophic perspective. Biochem. Syst. Ecol 2000, 28, 601–617. [Google Scholar]
- Brilli, F; Ciccioli, P; Frattoni, M; Prestininzi, M; Spanedda, AF; Loreto, F. Constitutive and herbivore-induced monoterpenes emitted by Populus euroamericana leaves are key volatiles that orient Chrysomela populi beetles. Plant Cell Environ 2009, 32, 542–552. [Google Scholar]
- Peñuelas, J; Llusià, J. The complexity of factors driving volatile organic compound emissions by plants. Biol. Plant 2001, 44, 481–487. [Google Scholar]
- Llusià, J; Peñuelas, J; Gimeno, BS. Seasonal and species-specific response of VOC emissions by Mediterranean woody plant to elevated ozone concentrations. Atmos. Environ 2002, 36, 3931–3938. [Google Scholar]
- Grote, R; Niinemets, U. Modeling volatile isoprenoid emissions–A story with split ends. Plant Biol 2008, 10, 8–28. [Google Scholar]
- Roselle, SJ. Effects of biogenic emission uncertainties on regional photochemical modelling of control strategies. Atmos. Environ 1994, 28, 1757–1772. [Google Scholar]
- Pinto, DM; Blande, JD; Nykanen, R; WenXia, D; Ner, AM; Holopainen, JK. Ozone degrades common herbivore-induced plant volatiles: Does this affect herbivore prey location by predators and parasitoids? J. Chem. Ecol 2007, 33, 683–694. [Google Scholar]
- Himanen, SJ; Nerg, AM; Nissinen, AM; Pinto, DM; Stewart, CN, Jr; Poppy, GM; Holopainen, JK. Effects of elevated carbon dioxide and ozone on volatile terpenoid emissions and multitrophic communication of transgenic insecticidal oilseed rape (Brassica napus.). New Phytol 2009, 181, 174–186. [Google Scholar]
- Freiwald, V; Haikio, E; Julkunen-Tiitto, R; Holopainen, JK; Oksanen, E. Elevated ozone modifies the feeding behaviour of the common leaf weevil on hybrid aspen through shifts in developmental, chemical, and structural properties of leaves. Entomol. Exp. Appl 2008, 128, 66–72. [Google Scholar]
- Alstad, DN; Edmunds, GF, Jr; Weinstein, LH. Effects of air pollutants on insect populations. Ann. Rev. Entomol 1982, 27, 369–384. [Google Scholar]
- Loreto, F; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products and reduces lipid peroxidation of cellular membrane. Plant Physiol 2001, 127, 1781–1787. [Google Scholar]
- Loreto, F; Ferranti, F; Mannozzi, M; Maris, C; Nascetti, C; Pasqualini, S. Ozone quenching properties of isoprene and its antioxidant role in plants. Plant Physiol 2001, 126, 993–1000. [Google Scholar]
- Velikova, V; Fares, S; Loreto, F. Isoprene and nitric oxide reduce damages in leaves exposed to oxidative stress. Plant Cell Environ 2008, 31, 1882–1894. [Google Scholar]
- Loreto, F; Pinelli, P; Manes, F; Kollist, K. Impact of ozone on monoterpene emission and evidence for an isoprene-like antioxidant action of monoterpenes emitted by Quercus ilex leaves. Tree Physiol 2004, 24, 361–367. [Google Scholar]
- Vitale, M; Salvatori, E; Loreto, F; Fares, S; Manes, F. Physiological responses of Quercus ilex to water stress and acute ozone exposure under controlled conditions. Water Air Soil Pollut 2008, 189, 113–125. [Google Scholar]
- Menser, HA; Chaplin, JF. Effects of ozone air pollution on nitrogen constituents, pH and water soluble ash of air-cured tobacco leaves. Tobacco Sci 1975, 19, 108–110. [Google Scholar]
- Aycock, MK. Phenotypic correlation of weather fleck with certain agronomic and chemical traits in Maryland tobacco. Tobacco Sci 1975, 19, 96–97. [Google Scholar]
- Jackson, DM; Rufty, TW; Heagle, AS; Severson, RF; Eckel, RVW. Survival and development of tobacco hornworm larvae on tobacco plants grown under elevated levels of ozone. J. Chem. Ecol 2000, 26, 1–19. [Google Scholar]
- Menser, HA; Chaplin, JF. Air pollution: Effects of phenol and alkaloid content of cured tobacco leaves. Tobacco Sci 1969, 13, 169–170. [Google Scholar]
- Jeffords, MR; Endress, AG. Possible role of ozone in tree defoliation by the gypsy moth (Lepidoptera: Lymantriidae). Environ. Entomol 1984, 13, 1249–1252. [Google Scholar]
- Endress, AG; Post, SC. Altered feeding preferences for Mexican bean beetle Epilachna varivestis for ozonated soybean foliage. Environ. Pollut 1985, 39, 9–16. [Google Scholar]
- Trumble, JT; Hare, JD; Musselmann, RC; McCool, PM. Ozone-induced changes in host-plant suitability: interactions of Keiferia lycopersicella and Lycopersicon esculentum. J. Chem. Ecol 1987, 13, 203–218. [Google Scholar]
- Chappelka, AH; Kraemer, ME; Mebrahtu, T; Rangappa, M; Benepal, PS. Effects of ozone on soybean resistance to Mexican bean beetle (Epilachna varivestis Mulsant). Environ. Exp. Bot 1988, 28, 53–60. [Google Scholar]
- Heagle, AS; Branderburg, RL; Burns, JC; Miller, JE. Ozone and carbon dioxide effects on spider mites in white clover and peanut. J. Environ. Qual 1994, 23, 1168–1176. [Google Scholar]
- Martin-Tanguy, J. Metabolism and function of polyamines in plants: Recentdevelopment (new approaches). Plant Growth Regul 2001, 34, 135–148. [Google Scholar]
- Walters, DR. Polyamines in plant-microbe interactions. Physiol. Mol. Plant Pathol 2000, 57, 137–146. [Google Scholar]
- Liu, J-H; Kitashiba, H; Wang, J; Ban, Y; Moriguchi, T. Polyamines and their ability to provide environmental stress tolerance to plants. Plant Biotechn 2007, 24, 117–126. [Google Scholar]
- Bagni, T; Tassoni, A. Biosynthesis oxidation and conjugation of aliphatic polyamines in higher plants. Amino Acids 2001, 20, 301–317. [Google Scholar]
- Rowland-Bamford, AJ; Borland, AM; Lea, PJ; Mansfield, TA. The role of arginine decarboxylase in modulating the sensitivity of barley to ozone. Environ. Pollut 1989, 61, 95–106. [Google Scholar]
- Langebartels, C; Wohlgemuth, H; Kschieschan, S; Grün, S; Sandermann, H. Oxidative burst and cell death in ozone-exposed plants. Plant Physiol. Biochem 2002, 40, 567–575. [Google Scholar]
- Ormrod, DP; Beckerson, DW. Polyamines as antiozonant for tomato. HortScience 1986, 21, 1070–1071. [Google Scholar]
- Bors, W; Langebartels, C; Michel, C; Sandermann, H. Polyamines as radical scavengers and protectans agaist ozone damages. Phytochemistry 1989, 28, 1589–1595. [Google Scholar]
- Bouchereau, A; Aziz, A; Larher, F; Martin-Tanguy, J. Polyamines and environmental challenges: Recent development. Plant Sci 1999, 140, 103–125. [Google Scholar]
- Hemantaranjan, A. Plant secondary metabolism in response to the environment. Adv. Plant Physiol 2008, 10, 31–89. [Google Scholar]
- Wink, M. Plant secondary metabolism:diversity, function and its evolution. Nat. Prod. Comm 2008, 3, 1205–1216. [Google Scholar]
- Violini, G. Ozone and plant pathogens: an overview. Riv. It. Pat. Veg 1995, 5, 113–130. [Google Scholar]
- Booker, F; Muntifering, R; McGrath, M; Burkey, K; Decoteau, D; Fiscus, E; Manning, W; Krupa, S; Chappelka, A; Grantz, D. The ozone component of global change: Potential effects on agricultural and horticultural plant yield, product quality and interactions with invasive species. J. Integrat. Plant Biol 2009, 51, 337–351. [Google Scholar]
- von Tiedemann, A; Firsching, KH. Interactive effects of elevated ozone and carbon dioxide on growth and yield of leaf rust-infected versus non-infected wheat. Environ. Pollut 2000, 72, 205–224. [Google Scholar]
- Lorenzini, G; Medeghini-Bonatti, P; Nali, C; Baroni Fornasiero, R. The protective ffect of rust infection against ozone, sulphur dioxide and paraquat toxicity symptoms in broad bean. Physiol. Mol. Plant Pathol 1994, 45, 263–279. [Google Scholar]
- Walters, DR; Boyle, C. Induced resistance and allocation costs: What is the impact of pathogen challenge? Physiol. Mol. Plant Pathol 2005, 66, 40–44. [Google Scholar]
- Walters, DR; Heil, M. Costs and trade offs associated with induced resistance. Physiol. Mol. Plant Pathol 2007, 71, 3–17. [Google Scholar]
- Iriti, M; Faoro, F. Bioactivity of grape chemicals for human health. Nat. Prod. Comm 2009, 4, 611–634. [Google Scholar]













© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Iriti, M.; Faoro, F. Chemical Diversity and Defence Metabolism: How Plants Cope with Pathogens and Ozone Pollution. Int. J. Mol. Sci. 2009, 10, 3371-3399. https://doi.org/10.3390/ijms10083371
Iriti M, Faoro F. Chemical Diversity and Defence Metabolism: How Plants Cope with Pathogens and Ozone Pollution. International Journal of Molecular Sciences. 2009; 10(8):3371-3399. https://doi.org/10.3390/ijms10083371
Chicago/Turabian StyleIriti, Marcello, and Franco Faoro. 2009. "Chemical Diversity and Defence Metabolism: How Plants Cope with Pathogens and Ozone Pollution" International Journal of Molecular Sciences 10, no. 8: 3371-3399. https://doi.org/10.3390/ijms10083371