A Microtube Array Membrane (MTAM) Encapsulated Live Fermenting Staphylococcus epidermidis as a Skin Probiotic Patch against Cutibacterium acnes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The S. epidermidis-encapsulated PSF MTAM
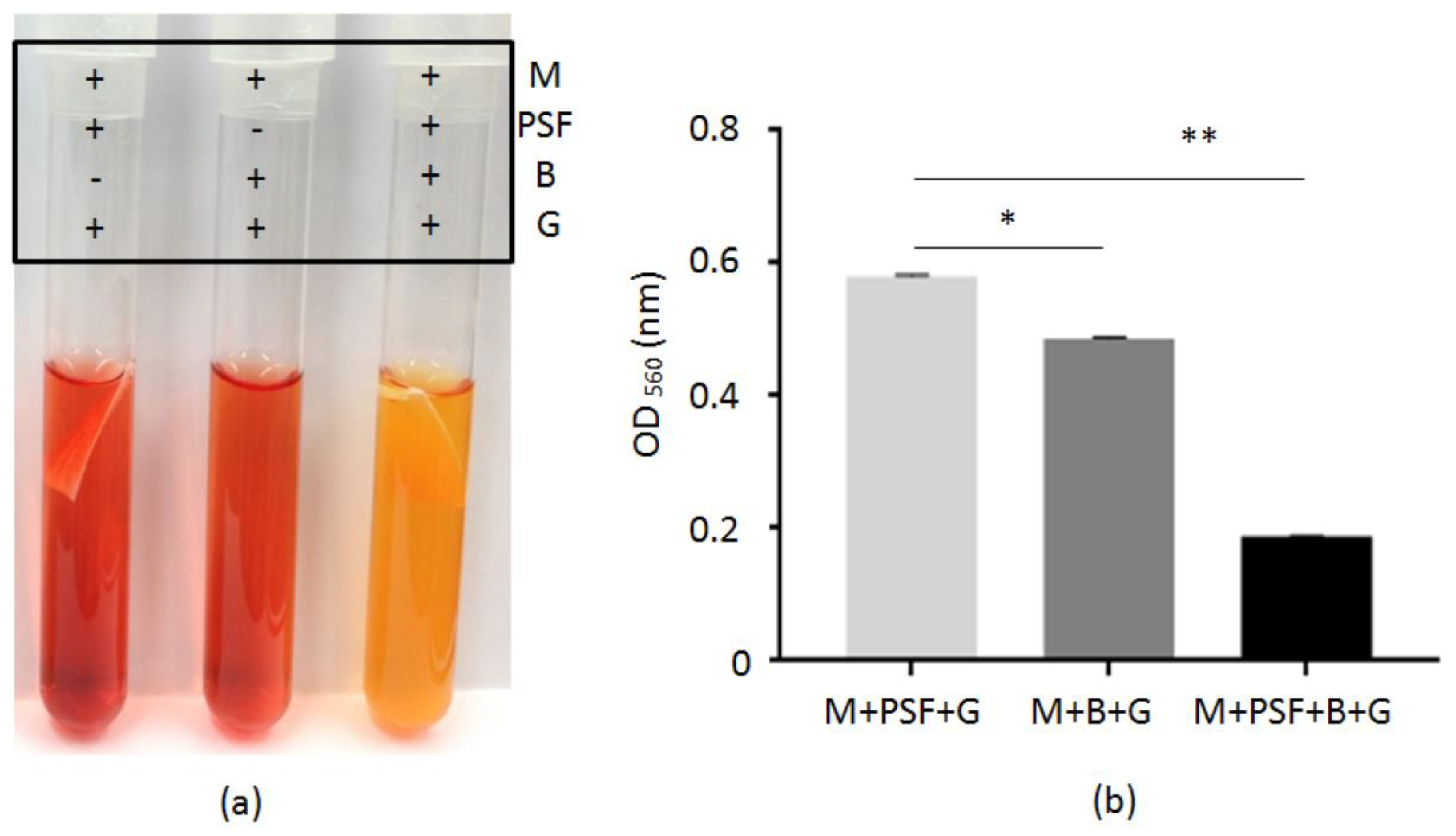
2.2. Glycerol Fermentation of S. epidermidis in PSF MTAM
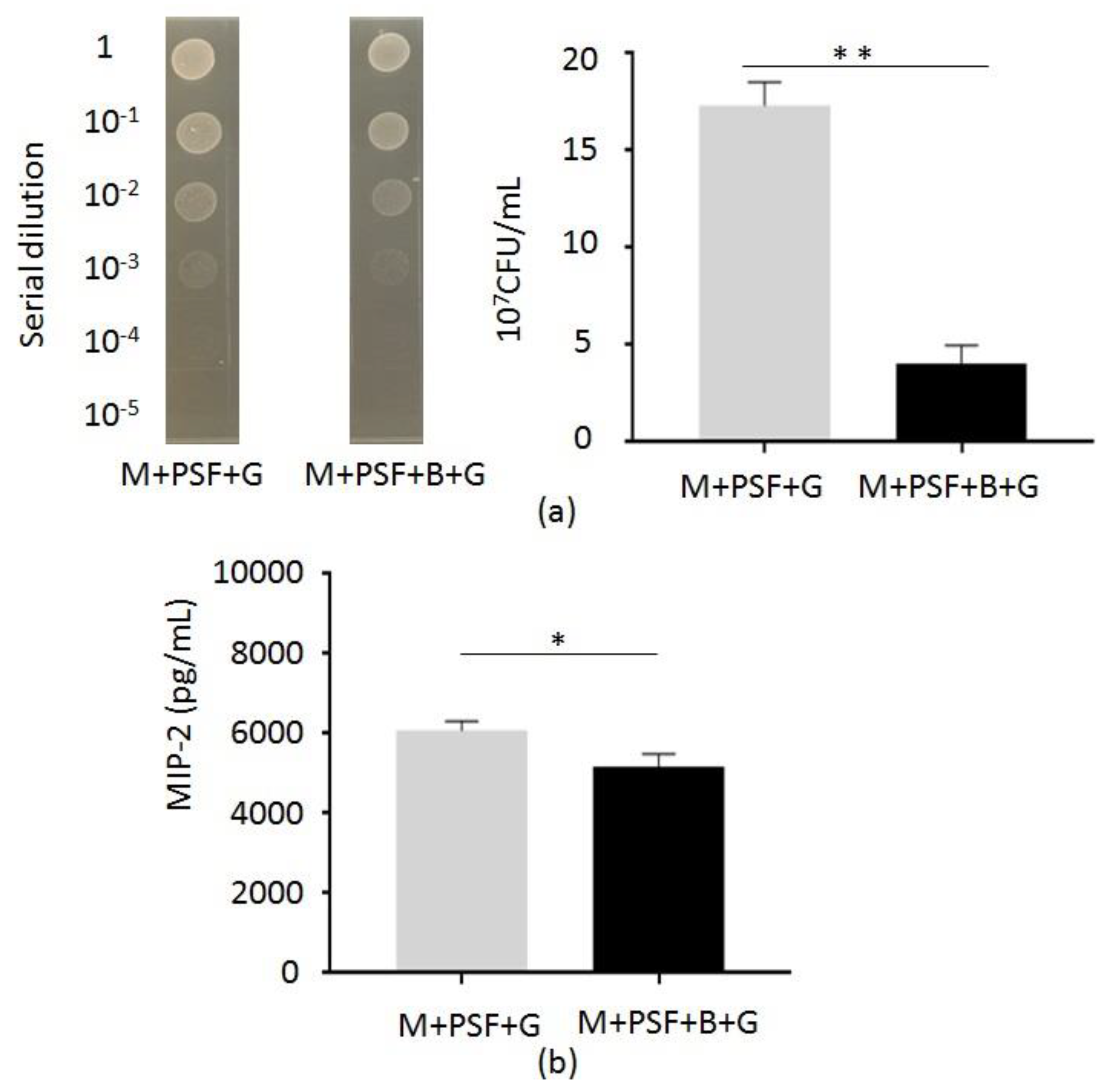
2.3. In Vivo Inhibition of C. acnes Growth and Inflammation by S. epidermidis Fermentation
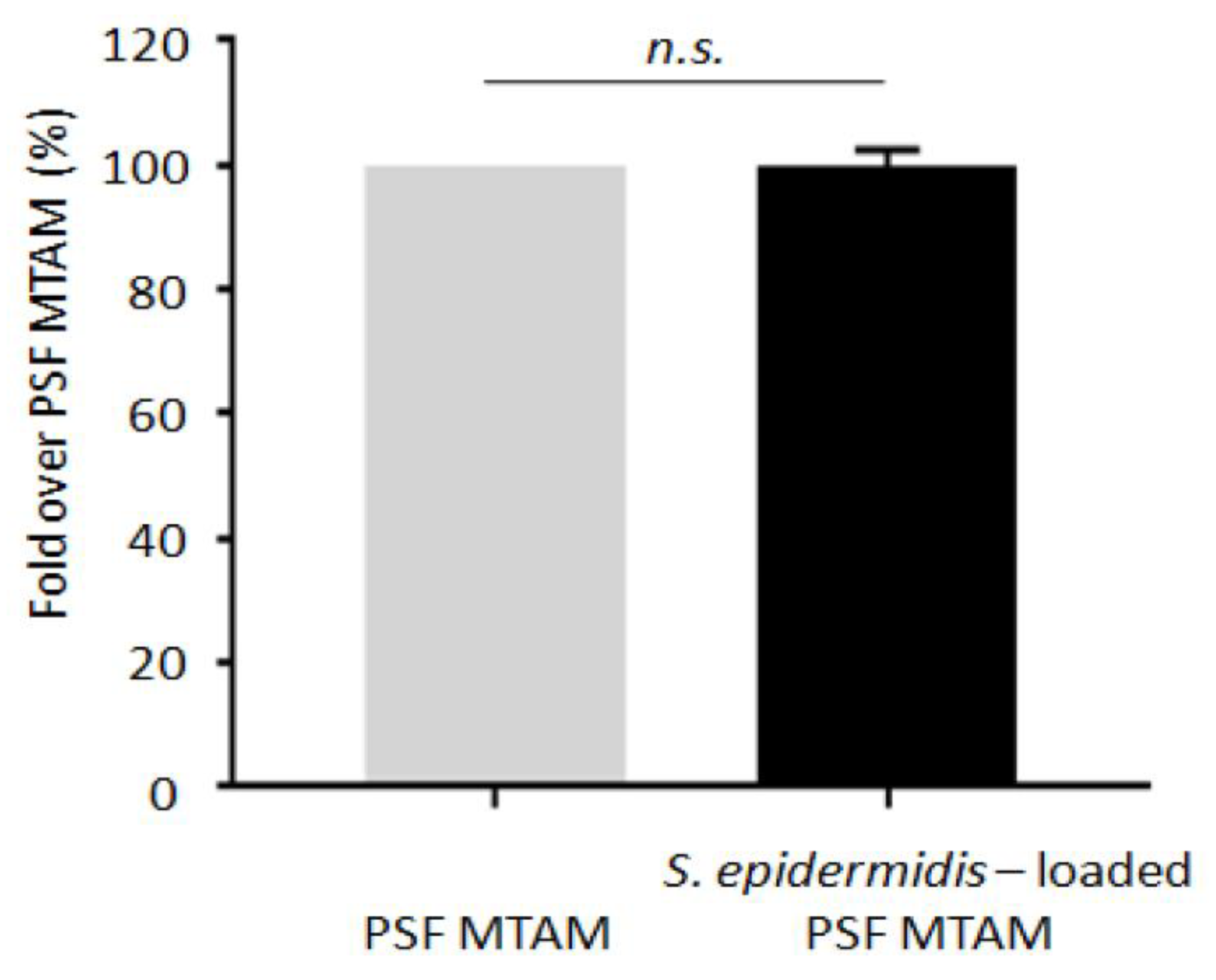
2.4. Detection of Bacterial Leaking from PSF MTAM
3. Discussion
4. Materials and Methods
4.1. Culture of Microorganisms
4.2. Fermentation of Bacteria
4.3. Fermentation of S. epidermidis in PSF MTAM Against C. acnes In Vivo
4.4. SEM
4.5. Bacterial Loads in Mouse Ears
4.6. ELISA
4.7. Detection of S. epidermidis Leaking from PSF MTAMs
4.8. Statistics
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244. [Google Scholar] [CrossRef] [PubMed]
- Bojar, R.A.; Holland, K.T. Acne and Propionibacterium acnes. Clin. Dermatol. 2004, 22, 375–379. [Google Scholar] [CrossRef]
- Liu, J.; Yan, R.; Zhong, Q.; Ngo, S.; Bangayan, N.J.; Nguyen, L.; Lui, T.; Liu, M.; Erfe, M.C.; Craft, N. The diversity and host interactions of Propionibacterium acnes bacteriophages on human skin. ISME J. 2015, 9, 2078. [Google Scholar] [CrossRef] [PubMed]
- MAYS, R.; Gordon, R.A.; Wilson, J.M.; Silapunt, S. New antibiotic therapies for acne and rosacea. Dermatol. Ther. 2012, 25, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Katsambas, A.D.; Stefanaki, C.; Cunliffe, W.J. Guidelines for treating acne. Clin. Dermatol. 2004, 22, 439–444. [Google Scholar] [CrossRef]
- Zaenglein, A.L.; Pathy, A.L.; Schlosser, B.J.; Alikhan, A.; Baldwin, H.E.; Berson, D.S.; Bowe, W.P.; Graber, E.M.; Harper, J.C.; Kang, S. Guidelines of care for the management of acne vulgaris. J. Am. Acad. Dermatol. 2016, 74, 945–973. [Google Scholar] [CrossRef]
- Hull, P.R.; D’Arcy, C. Isotretinoin use and subsequent depression and suicide. Am. J. Clin. Dermatol. 2003, 4, 493–505. [Google Scholar] [CrossRef]
- Wang, Y.; Kuo, S.; Shu, M.; Yu, J.; Huang, S.; Dai, A.; Gallo, R.L.; Huang, C.-M. Staphylococcus epidermidis in the human skin microbiome mediates fermentation to inhibit the growth of Propionibacterium acnes: Implications of probiotics in acne vulgaris. Appl. Microbiol. Biotechnol. 2014, 98, 411–424. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Yu, J.; Huang, S.; Wang, Z.; Chun, K.A.; Lee, T.L.; Chen, Y.-T.; Gallo, R.L.; Huang, C.-M. A Co-Drug of Butyric Acid Derived from Fermentation Metabolites of the Human Skin Microbiome Stimulates Adipogenic Differentiation of Adipose-Derived Stem Cells: Implications in Tissue Augmentation. J. Investig. Dermatol. 2017, 137, 46–56. [Google Scholar] [CrossRef]
- Christensen, G.J.; Scholz, C.F.; Enghild, J.; Rohde, H.; Kilian, M.; Thürmer, A.; Brzuszkiewicz, E.; Lomholt, H.B.; Brüggemann, H. Antagonism between Staphylococcus epidermidis and Propionibacterium acnes and its genomic basis. BMC Genom. 2016, 17, 152. [Google Scholar] [CrossRef]
- Rajiv, P.; Nitesh, K.; Raj, K.; Hemant, G.K. Staphylococcus epidermidis in human skin microbiome associated with acne: A cause of disease or defence? Res. J. Biotechnol. 2013, 8, 78–82. [Google Scholar]
- Wang, Y.; Kao, M.-S.; Yu, J.; Huang, S.; Marito, S.; Gallo, R.L.; Huang, C.-M. A precision microbiome approach using sucrose for selective augmentation of Staphylococcus epidermidis fermentation against Propionibacterium acnes. Int. J. Mol. Sci. 2016, 17, 1870. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, F.A. Development of probiotics as biologic drugs. Clin. Infect. Dis. 2008, 46, S125–S127. [Google Scholar] [CrossRef] [PubMed]
- Frenot, A.; Chronakis, I.S. Polymer nanofibers assembled by electrospinning. Curr. Opin. Colloid Interface Sci. 2003, 8, 64–75. [Google Scholar] [CrossRef]
- Wendorff, J.H.; Agarwal, S.; Greiner, A. Electrospinning: Materials, Processing, and Applications; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Jang, J.-S.; Cho, Y.; Jeong, G.-T.; Kim, S.-K. Optimization of saccharification and ethanol production by simultaneous saccharification and fermentation (SSF) from seaweed, Saccharina japonica. Bioprocess Biosyst. Eng. 2012, 35, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.C.; Lee, S.Y.; Tseng, W.C.; Shu, Y.C.; Lu, J.C.; Shie, H.S.; Chen, C.C. Formation of Highly Aligned, Single-Layered, Hollow Fibrous Assemblies and the Fabrication of Large Pieces of PLLA Membranes. Macromol. Mater. Eng. 2012, 297, 115–122. [Google Scholar] [CrossRef]
- Chen, C.-C.; Wu, C.-H.; Wu, J.-J.; Chiu, C.-C.; Wong, C.-H.; Tsai, M.-L.; Lin, H.-T.V. Accelerated bioethanol fermentation by using a novel yeast immobilization technique: Microtube array membrane. Process Biochem. 2015, 50, 1509–1515. [Google Scholar] [CrossRef]
- Frank, D.N.; Feazel, L.M.; Bessesen, M.T.; Price, C.S.; Janoff, E.N.; Pace, N.R. The human nasal microbiota and Staphylococcus aureus carriage. PLoS ONE 2010, 5, e10598. [Google Scholar] [CrossRef]
- Ji, G.; Beavis, R.; Novick, R.P. Bacterial interference caused by autoinducing peptide variants. Science 1997, 276, 2027–2030. [Google Scholar] [CrossRef]
- Nicoll, T.; Jensen, M. Staphylococcosis of turkeys. 5. Large-scale control programs using bacterial interference. Avian Dis. 1987, 31, 85–88. [Google Scholar] [CrossRef]
- Wei, W.; Cao, Z.; Zhu, Y.-L.; Wang, X.; Ding, G.; Xu, H.; Jia, P.; Qu, D.; Danchin, A.; Li, Y. Conserved genes in a path from commensalism to pathogenicity: Comparative phylogenetic profiles of Staphylococcus epidermidis RP62A and ATCC12228. BMC Genom. 2006, 7, 112. [Google Scholar] [CrossRef]
- Whitehead, S.; Leavitt, R.; Jensen, M. Staphylococcosis of turkeys. 6. Development of penicillin resistance in an interfering strain of Staphylococcus epidermidis. Avian Dis. 1993, 37, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.; Jensen, M. Staphylococcosis of turkeys. 4. Characterization of a bacteriocin produced by an interfering Staphylococcus. Avian Dis. 1987, 31, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Dimarzio, L.; Cinque, B.; Cupelli, F.; De Simone, C.; Cifone, M.; Giuliani, M. Increase of skin-ceramide levels in aged subjects following a short-term topical application of bacterial sphingomyelinase from Streptococcus thermophilus. Int. J. Immunopathol. Pharmacol. 2008, 21, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Gueniche, A.; Benyacoub, J.; Philippe, D.; Bastien, P.; Kusy, N.; Breton, L.; Blum, S.; Castiel-Higounenc, I. Lactobacillus paracasei CNCM I-2116 (ST11) inhibits substance P-induced skin inflammation and accelerates skin barrier function recovery in vitro. Eur. J. Dermatol. 2010, 20, 731–737. [Google Scholar]
- Kourkoutas, Y.; Bekatorou, A.; Banat, I.M.; Marchant, R.; Koutinas, A. Immobilization technologies and support materials suitable in alcohol beverages production: A review. Food Microbiol. 2004, 21, 377–397. [Google Scholar] [CrossRef]
- Lee, K.H.; Choi, I.S.; Kim, Y.-G.; Yang, D.-J.; Bae, H.-J. Enhanced production of bioethanol and ultrastructural characteristics of reused Saccharomyces cerevisiae immobilized calcium alginate beads. Bioresource Technol. 2011, 102, 8191–8198. [Google Scholar] [CrossRef]
- Chandel, A.K.; Narasu, M.L.; Chandrasekhar, G.; Manikyam, A.; Rao, L.V. Use of Saccharum spontaneum (wild sugarcane) as biomaterial for cell immobilization and modulated ethanol production by thermotolerant Saccharomyces cerevisiae VS3. Bioresource Technol. 2009, 100, 2404–2410. [Google Scholar] [CrossRef]
- Eiadpum, A.; Limtong, S.; Phisalaphong, M. High-temperature ethanol fermentation by immobilized coculture of Kluyveromyces marxianus and Saccharomyces cerevisiae. J. Biosci. Bioeng. 2012, 114, 325–329. [Google Scholar] [CrossRef]
- Levine, R.M.; Rasmussen, J.E. Intralesional corticosteroids in the treatment of nodulocystic acne. Arch. Dermatol. 1983, 119, 480–481. [Google Scholar] [CrossRef]
- Al-Kathiri, L.; Al-Najjar, T. Severe Nodulocystic Acne not Responding to Isotretinoin Therapy Successfully Treated with Oral Dapsone. Oman Med. J. 2018, 33, 43. [Google Scholar] [CrossRef] [PubMed]
- Fluhr, J.; Darlenski, R.; Surber, C. Glycerol and the skin: Holistic approach to its origin and functions. Br. J. Dermatol. 2008, 159, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Yoneya, T.; Nishijima, Y. Determination of free glycerol on human skin surface. Biol. Mass Spectrom. 1979, 6, 191–193. [Google Scholar] [CrossRef]
- Ricke, S. Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult. Sci. 2003, 82, 632–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, M.; Wang, Y.; Yu, J.; Kuo, S.; Coda, A.; Jiang, Y.; Gallo, R.L.; Huang, C.-M. Fermentation of Propionibacterium acnes, a commensal bacterium in the human skin microbiome, as skin probiotics against methicillin-resistant Staphylococcus aureus. PLoS ONE 2013, 8, e55380. [Google Scholar] [CrossRef] [PubMed]
- Kligman, A.M. An overview of acne. J. Investig. Dermatol. 1974, 62, 268–287. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.-F.; Nakatsuji, T.; Zhu, W.; Gallo, R.L.; Huang, C.-M. Passive immunoprotection targeting a secreted CAMP factor of Propionibacterium acnes as a novel immunotherapeutic for acne vulgaris. Vaccine 2011, 29, 3230–3238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valanne, S.; McDowell, A.; Ramage, G.; Tunney, M.M.; Einarsson, G.G.; O’Hagan, S.; Wisdom, G.B.; Fairley, D.; Bhatia, A.; Maisonneuve, J.-F. CAMP factor homologues in Propionibacterium acnes: A new protein family differentially expressed by types I and II. Microbiology 2005, 151, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Han, Q.; Chen, G.; Huang, Y.; Zhao, L.-X.; Berta, T.; Gao, Y.-J.; Ji, R.-R. Toll-like receptor 4 contributes to chronic itch, alloknesis and spinal astrocyte activation in male mice. Pain 2016, 157, 806. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hata, T.R.; Tong, Y.L.; Kao, M.-S.; Zouboulis, C.C.; Gallo, R.L.; Huang, C.-M. The anti-inflammatory activities of Propionibacterium acnes CAMP factor-targeted acne vaccines. J. Investig. Dermatol. 2018, 138, 2355–2364. [Google Scholar] [CrossRef] [PubMed]
- Burtenshaw, J. The mechanism of self-disinfection of the human skin and its appendages. Epidemiol. Infect. 1942, 42, 184–210. [Google Scholar] [CrossRef]
- Garland, S. Short chain fatty acids may elicit an innate immune response from preadipocytes: A potential link between bacterial infection and inflammatory diseases. Med. Hypotheses 2011, 76, 881–883. [Google Scholar] [CrossRef] [PubMed]
- Schröder, O.; Opritz, J.; Stein, J. Substrate and inhibitor specificity of butyrate uptake in apical membrane vesicles of the rat distal colon. Digestion 2000, 62, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Stein, J.; Zores, M.; Schröder, O. Short-chain fatty acid (SCFA) uptake into Caco-2 cells by a pH-dependent and carrier mediated transport mechanism. Eur. J. Nutr. 2000, 39, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Hobdy, E.; Murren, J. AN-9 (Titan). Curr. Opin. Investig. Drugs 2004, 5, 628–634. [Google Scholar] [PubMed]
- Mondragón, M.; Trujillo, G.; Moggio, I.; Arias, E. Luminescent polylactic acid and polysulfone electrospun fibers containing europium (III) complexes. Eur. Polym. J. 2016, 80, 126–133. [Google Scholar] [CrossRef]
- Berins, M. Plastics Engineering Handbook of the Society of the Plastics Industry; Springer Science & Business Media: Berlin, Germany, 1991. [Google Scholar]
- Brydson, J.A. Plastics Materials; Elsevier: Amsterdam, The Netherlands, 1999. [Google Scholar]
- Harper, C.A. Modern Plastics Handbook: Handbook; McGraw-Hill Professional: New York, NY, USA, 2000. [Google Scholar]
- Yu, X.; Liu, F.; Wang, L.; Xiong, Z.; Wang, Y. Robust poly (lactic acid) membranes improved by polysulfone-g-poly (lactic acid) copolymers for hemodialysis. RSC Adv. 2015, 5, 78306–78314. [Google Scholar] [CrossRef]
- Crompton, T.R. Physical Testing of Plastics; Smithers Rapra Technology: Shrewsbury, UK, 2012. [Google Scholar]
- Chew, C.; Wu, C.; Chen, C. A novel electrospun Microtube Array Membrane (MTAM) based low cost conceptual tubular Microbial Fuel Cell (MFC). Eur. Polym. J. 2016, 83, 138–147. [Google Scholar] [CrossRef]
- Mastronardi, C.C.; Ramírez-Arcos, S. Quantitative PCR for detection and discrimination of the bloodborne pathogen Staphylococcus epidermidis in platelet preparations using divIVA and icaA as target genes. Can. J. Microbiol. 2007, 53, 1222–1231. [Google Scholar] [CrossRef]
- Hardikar, A.A.; Farr, R.J.; Joglekar, M.V. Circulating microRNAs: Understanding the limits for quantitative measurement by real-time PCR. J. Am. Heart Assoc. 2014, 3, e000792. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, A.-J.; Marito, S.; Yang, J.-J.; Keshari, S.; Chew, C.-H.; Chen, C.-C.; Huang, C.-M. A Microtube Array Membrane (MTAM) Encapsulated Live Fermenting Staphylococcus epidermidis as a Skin Probiotic Patch against Cutibacterium acnes. Int. J. Mol. Sci. 2019, 20, 14. https://doi.org/10.3390/ijms20010014
Yang A-J, Marito S, Yang J-J, Keshari S, Chew C-H, Chen C-C, Huang C-M. A Microtube Array Membrane (MTAM) Encapsulated Live Fermenting Staphylococcus epidermidis as a Skin Probiotic Patch against Cutibacterium acnes. International Journal of Molecular Sciences. 2019; 20(1):14. https://doi.org/10.3390/ijms20010014
Chicago/Turabian StyleYang, Albert-Jackson, Shinta Marito, John-Jackson Yang, Sunita Keshari, Chee-Ho Chew, Chien-Chung Chen, and Chun-Ming Huang. 2019. "A Microtube Array Membrane (MTAM) Encapsulated Live Fermenting Staphylococcus epidermidis as a Skin Probiotic Patch against Cutibacterium acnes" International Journal of Molecular Sciences 20, no. 1: 14. https://doi.org/10.3390/ijms20010014