Phytotoxicity, Bioaccumulation, and Degradation of Nonylphenol in Different Microalgal Species without Bacterial Influences

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
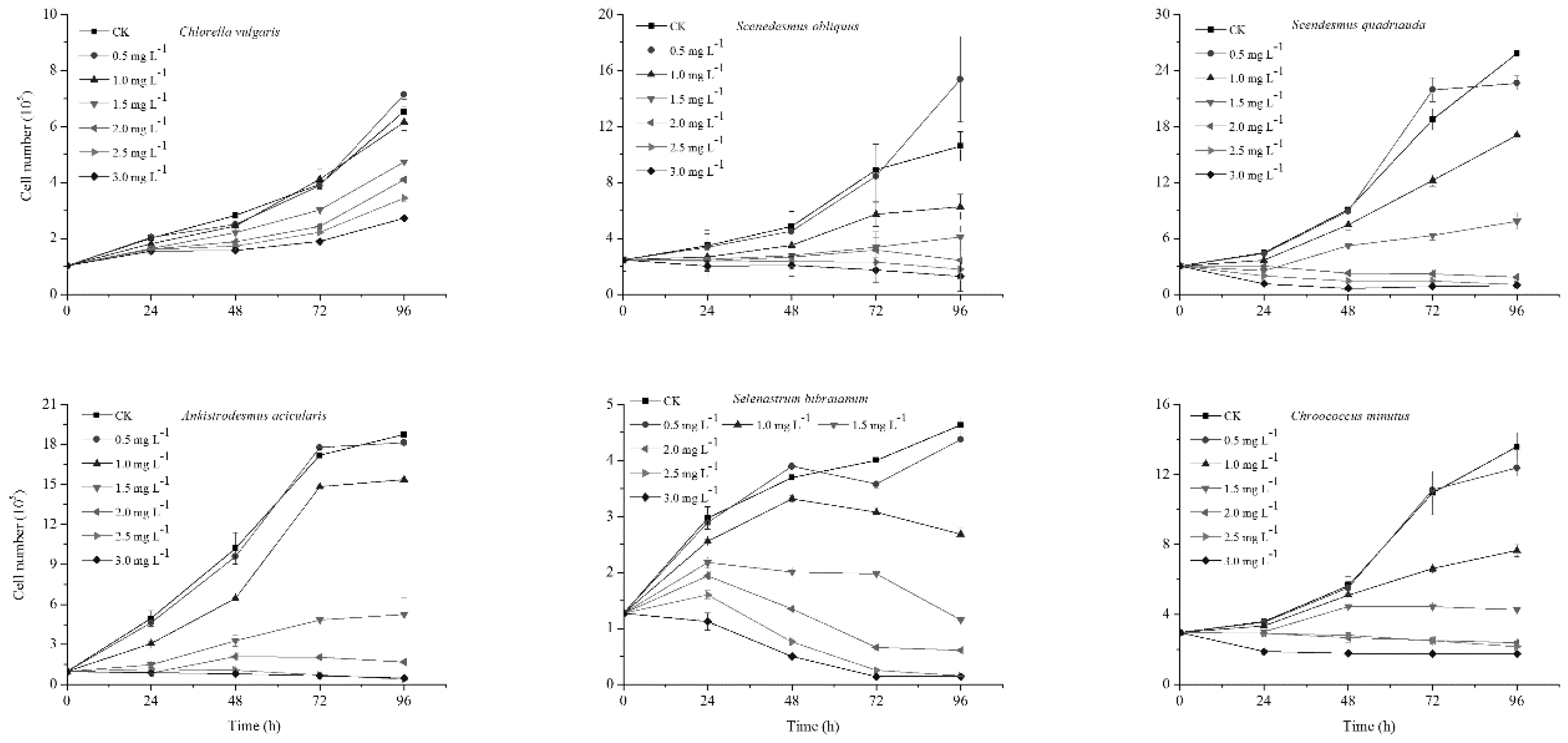
2.1. Affect of NP on Algal Growth
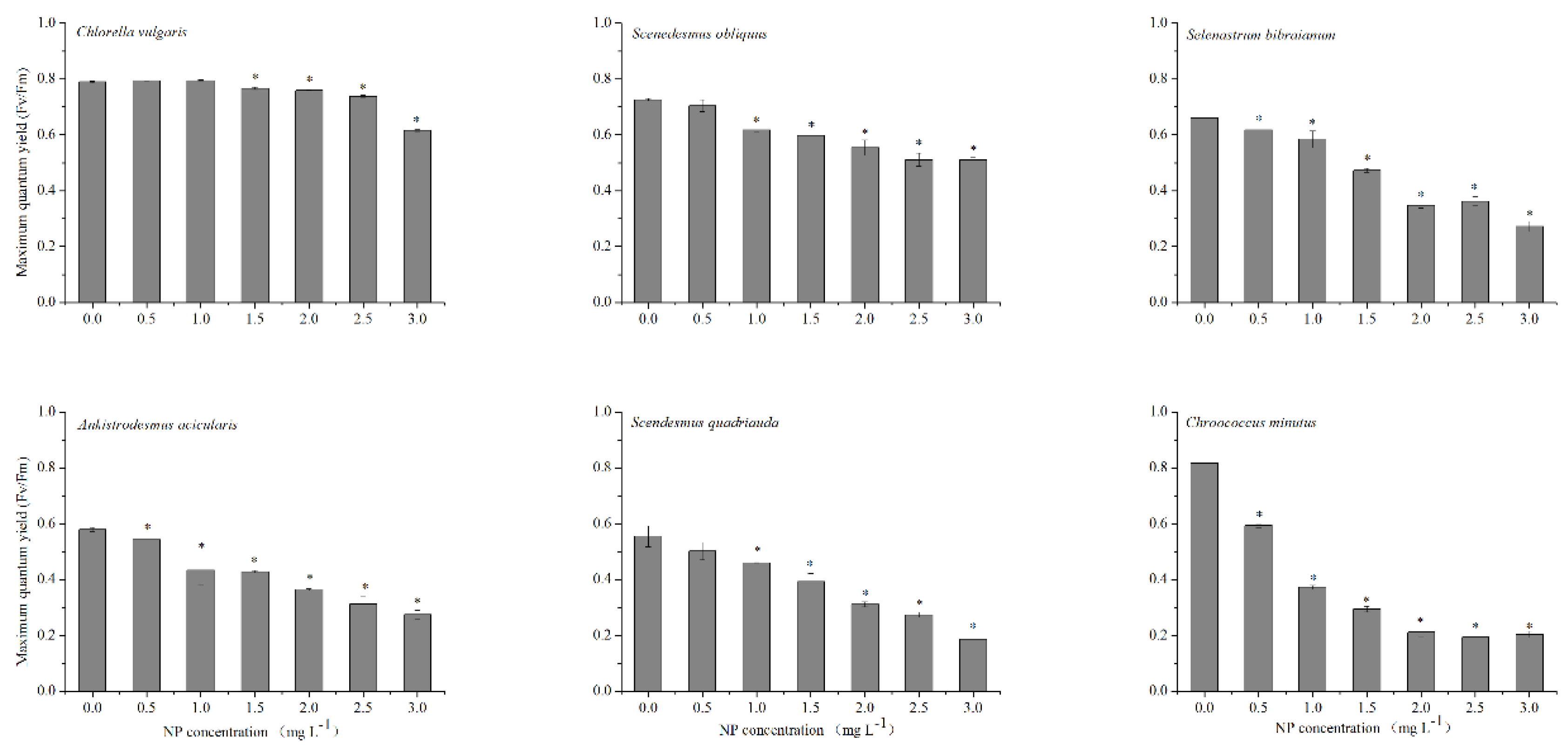
2.2. Influence of NP on Chlorophyll Fluorescence
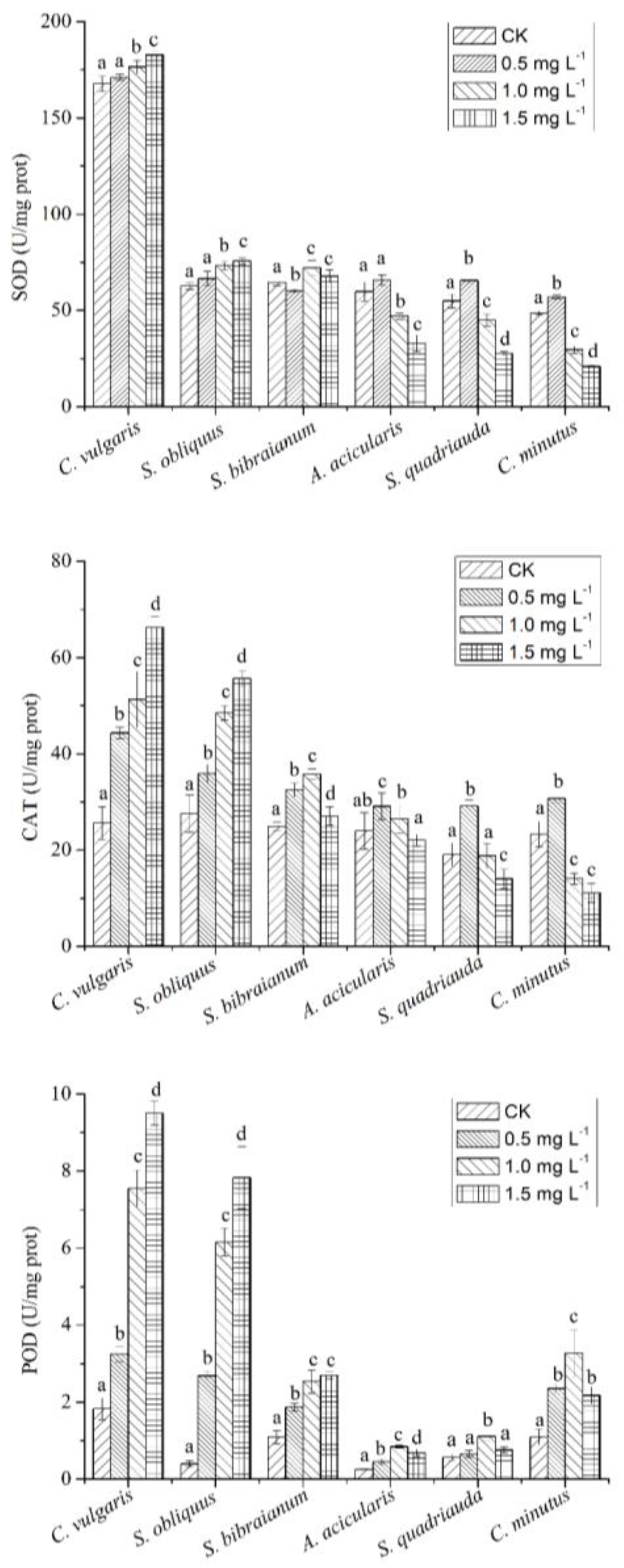
2.3. The Relationship between NP and Antioxidant Enzymes
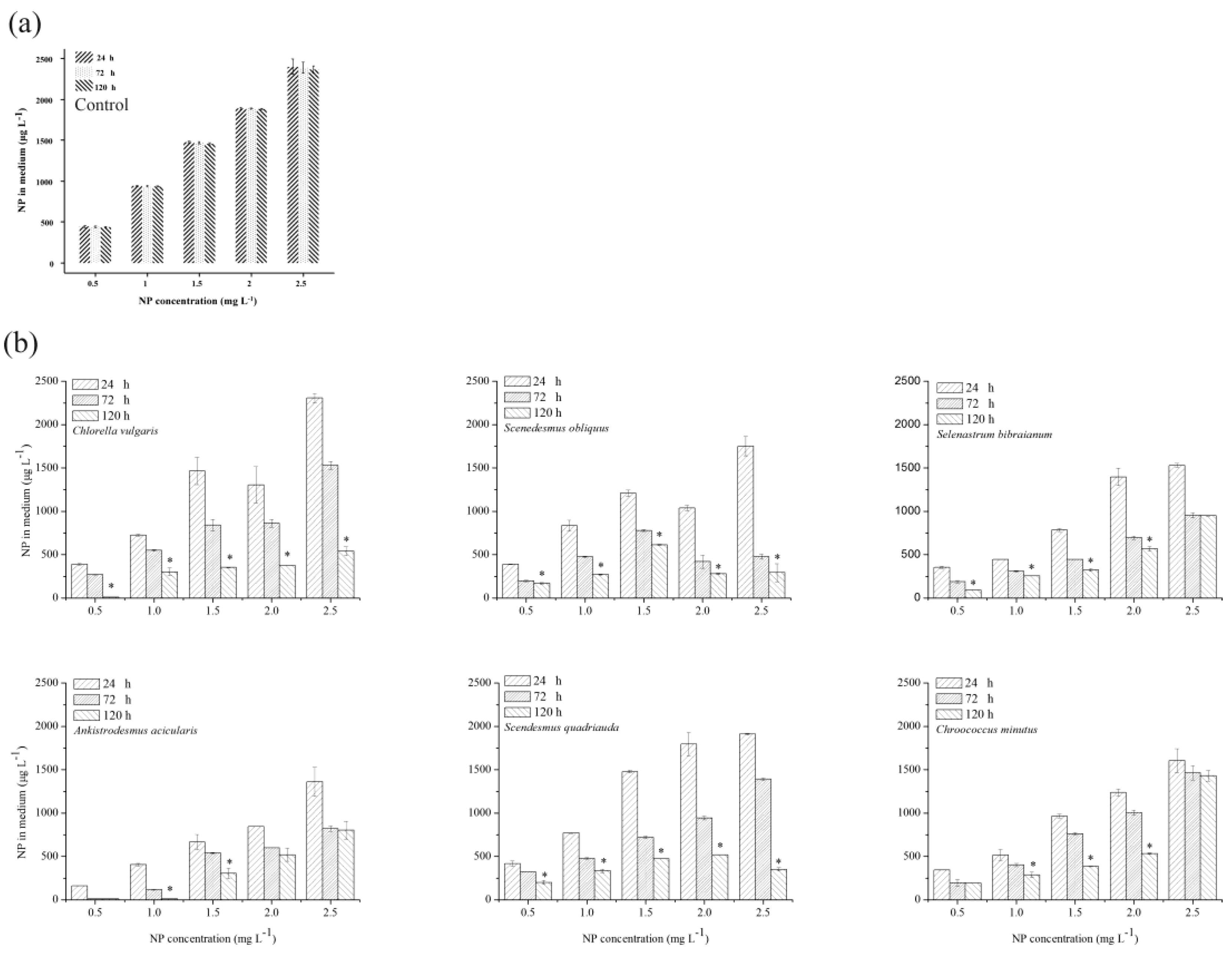
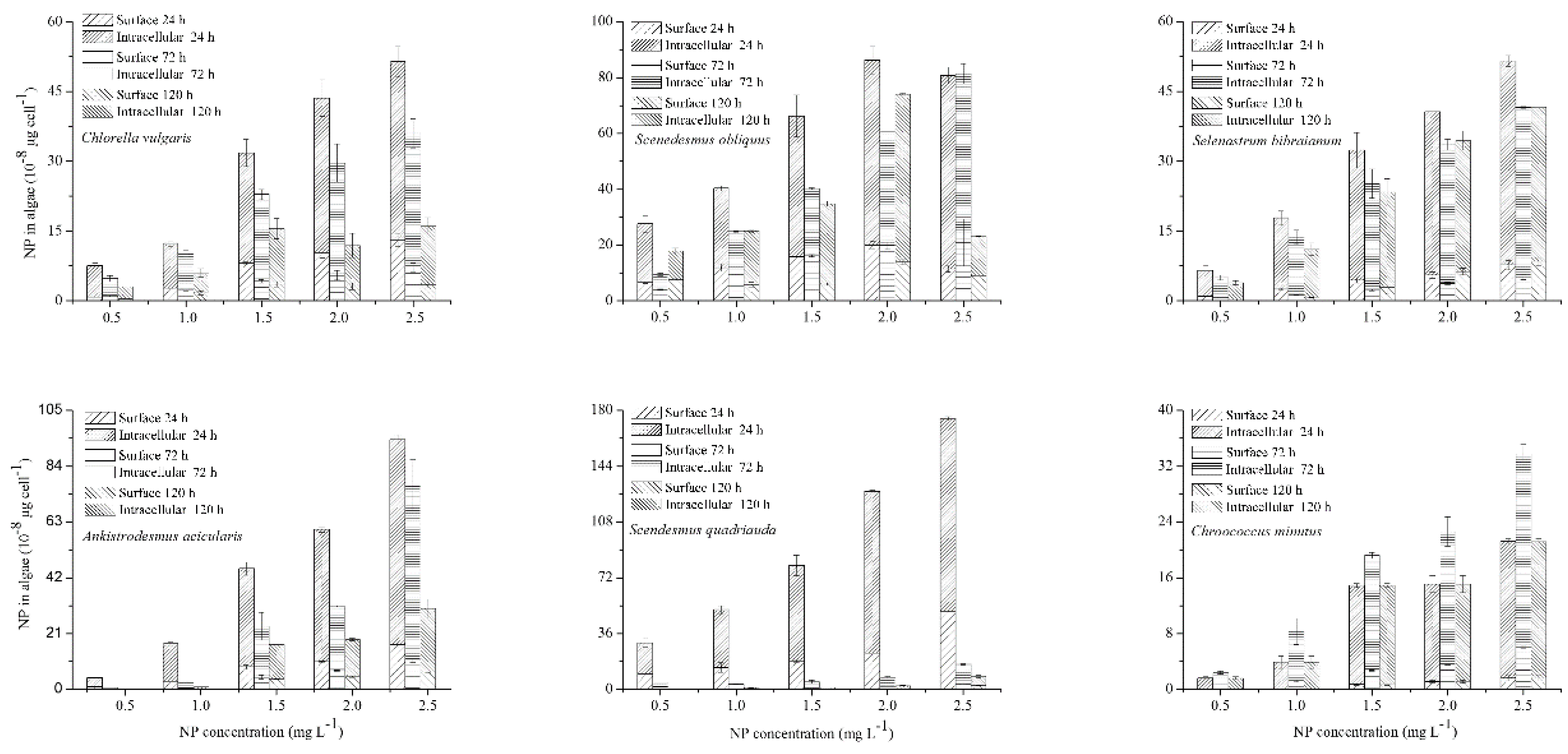
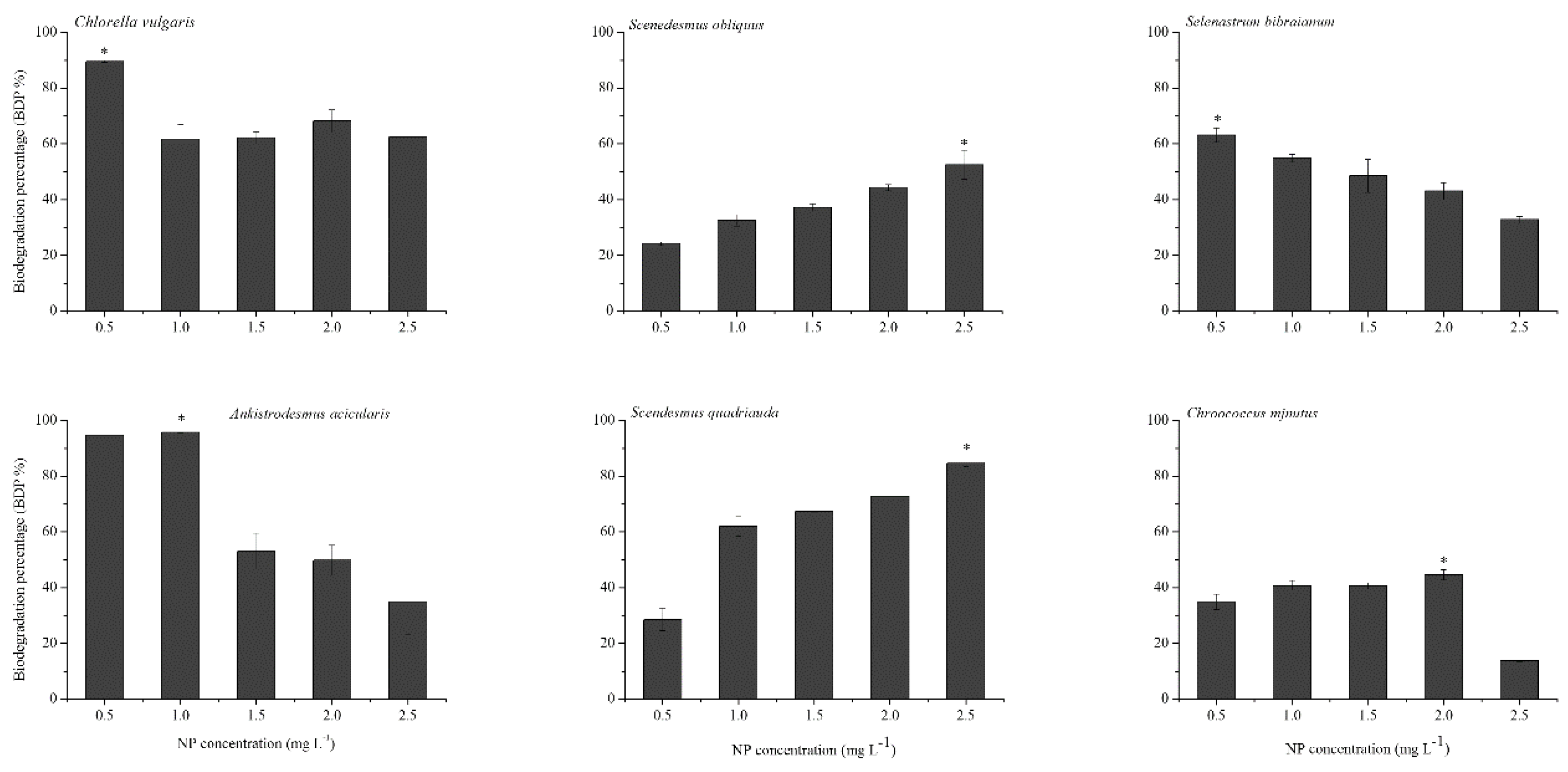
2.4. NP Accumulation and Degradation in Algae
3. Materials and Methods
3.1. Reagents
3.2. Algal Culture and Treatment
3.3. Removal of Bacteria from Algal Cultures
3.4. Nonylphenol Treatments
3.5. Determination of Algal Growth and Inhibitory Effect Concentration
3.6. Measurement of Fluorescence Transient
3.7. Assay of Antioxidant Enzyme Activity
3.8. Quantification of NP in Culture Medium and Algae
3.9. Determination of Biodegradation Percentage of NP
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sood, A.; Uniyal, P.L.; Prasanna, R.; Ahluwalia, A.S. Phytoremediation Potential of Aquatic Macrophyte, Azolla. AMBIO 2012, 41, 122–137. [Google Scholar] [CrossRef] [Green Version]
- Soares, A.; Guieysse, B.; Jefferson, B.; Cartmell, E.; Lester, J.N. Nonylphenol in the environment: A critical review on occurrence, fate, toxicity and treatment in wastewaters. Environ. Int. 2008, 34, 1033–1049. [Google Scholar] [CrossRef]
- Commission, E. Directive 2003/53/EC of the European Parliament and of the Council of 18 June 2003 amending for the 26th time Council Directive 76/769/EEC relating to restrictions on the marketing and use of certain dangerous substances and preparations (nonylphenol, nonylphenol ethoxylate and cement). In Official Journal of the European Union; Publications Office of the European Union: Luxembourg, 2003; pp. 24–178. [Google Scholar]
- Zgoła-Grześkowiak, A. Dispersive liquid–liquid microextraction applied to isolation and concentration of alkylphenols and their short-chained ethoxylates in water samples. J. Chromatogr. A 2010, 1217, 1761–1766. [Google Scholar] [CrossRef]
- Mao, Z.; Zheng, X.F.; Zhang, Y.-Q.; Tao, X.-X.; Li, Y.; Wang, W. Occurrence and Biodegradation of Nonylphenol in the Environment. Int. J. Mol. Sci. 2012, 13, 491–505. [Google Scholar] [CrossRef]
- Klečka, G.M.; Naylor, C.G.; Staples, C.A.; Losey, B. Occurrence of Nonylphenol Ethoxylates and Their Metabolites in Municipal Wastewater Treatment Plants and Receiving Waters. Water Environ. Res. 2010, 82, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Hu, J.; An, W.; Yang, M. Detection and Occurrence of Chlorinated Byproducts of Bisphenol A, Nonylphenol, and Estrogens in Drinking Water of China: Comparison to the Parent Compounds. Environ. Sci. Technol. 2013, 47, 10841–10850. [Google Scholar] [CrossRef] [PubMed]
- Newsted, J.L. Effect of light, temperature, and pH on the accumulation of phenol by Selenastrum capricornutum, a green alga. Ecotoxicol. Environ. Saf. 2004, 59, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Dosnon-Olette, R.; Trotel-Aziz, P.; Couderchet, M.; Eullaffroy, P. Fungicides and herbicide removal in Scenedesmus cell suspensions. Chemosphere 2010, 79, 117–123. [Google Scholar] [CrossRef] [PubMed]
- McCormick, P.V.; Cairns, J. Algae as indicators of environmental change. J Appl Phycol 1994, 6, 509–526. [Google Scholar] [CrossRef]
- Eullaffroy, P.; Vernet, G. The F684/F735 chlorophyll fluorescence ratio: A potential tool for rapid detection and determination of herbicide phytotoxicity in algae. Water. Res. 2003, 37, 1983–1990. [Google Scholar] [CrossRef]
- Corcoll, N.; Bonet, B.; Leira, M.; Guasch, H. Chl-a fluorescence parameters as biomarkers of metal toxicity in fluvial biofilms: An experimental study. Hydrobiologia 2011, 673, 119–136. [Google Scholar] [CrossRef]
- Sun, X.; Zhong, Y.; Huang, Z.; Yang, Y. Selenium Accumulation in Unicellular Green Alga Chlorella vulgaris and Its Effects on Antioxidant Enzymes and Content of Photosynthetic Pigments. PLoS ONE 2014, 9, e112270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvíderová, J. Rapid algal toxicity assay using variable chlorophyll fluorescence for Chlorella kessleri (chlorophyta). Environ. Toxicol 2010, 25, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.J.; Berges, J.A.; Young, E.B. Rapid effects of diverse toxic water pollutants on chlorophyll a fluorescence: Variable responses among freshwater microalgae. Water. Res. 2012, 46, 2615–2626. [Google Scholar] [CrossRef]
- Wang, S.; Chen, F.; Mu, S.; Zhang, D.; Pan, X.; Lee, D.-J. Simultaneous analysis of photosystem responses of Microcystis aeruginoga under chromium stress. Ecotox. Environ. Safe 2013, 88, 163–168. [Google Scholar] [CrossRef]
- Wang, J.; Xie, P. Antioxidant enzyme activities of Microcystis aeruginosa in response to nonylphenols and degradation of nonylphenols by M. aeruginosa. Environ. Geochem. Health. 2007, 29, 375–383. [Google Scholar] [CrossRef]
- Shan, J.; Jiang, B.; Yu, B.; Li, C.; Sun, Y.; Guo, H.; Wu, J.; Klumpp, E.; Schaeffer, A.; Ji, R. Isomer-Specific Degradation of Branched and Linear 4-Nonylphenol Isomers in an Oxic Soil. Environ. Sci. Technol. 2011, 45, 8283–8289. [Google Scholar] [CrossRef]
- Xu, P.; Lai, C.; Zeng, G.; Huang, D.; Chen, M.; Song, B.; Peng, X.; Wan, J.; Hu, L.; Duan, A.; et al. Enhanced bioremediation of 4-nonylphenol and cadmium co-contaminated sediment by composting with Phanerochaete chrysosporium inocula. Bioresour. Technol. 2018, 250, 625–634. [Google Scholar] [CrossRef]
- He, N.; Sun, X.; Zhong, Y.; Sun, K.; Liu, W.; Duan, S. Removal and Biodegradation of Nonylphenol by Four Freshwater Microalgae. Int. J. Environ. Res. Public Health 2016, 13, 1239. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Zhou, P. Advance in the studies on effect of environmental organic pollutants on the algae growth. Acta Hydrobiol. Sin. 2002, 26, 529–535. [Google Scholar]
- Tsang, C.K.; Lau, P.S.; Tam, N.F.Y.; Wong, Y.S. Biodegradation capacity of tributyltin by two Chlorella species. Environ. Pollut. 1999, 105, 289–297. [Google Scholar] [CrossRef]
- Andrade, L.R.; Leal, R.N.; Noseda, M.; Duarte, M.E.R.; Pereira, M.S.; Mourão, P.A.S.; Farina, M.; Amado Filho, G.M. Brown algae overproduce cell wall polysaccharides as a protection mechanism against the heavy metal toxicity. Mar. Pollut. Bull. 2010, 60, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Osundeko, O.; Davies, H.; Pittman, J.K. Oxidative stress-tolerant microalgae strains are highly efficient for biofuel feedstock production on wastewater. Biomass Bioenergy 2013, 56, 284–294. [Google Scholar] [CrossRef]
- Sutherland, D.; Ralph, P. Microalgal bioremediation of emerging contaminants - Opportunities and challenges. Water. Res. 2019, 164, 114921. [Google Scholar] [CrossRef]
- Servos, M. Review of the Aquatic Toxicity, Estrogenic Responses and Bioaccumulation of Alkylphenols and Alkylphenol Polyethoxylates. Water Qual. Res. J. Can. 1999, 34, 123–177. [Google Scholar] [CrossRef]
- Wang, J.; Xie, P.; Guo, N. Effects of nonylphenol on the growth and microcystin production of Microcystis strains. Environ. Res. 2007, 103, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Qian, X.J.; An, M.; Duan, S.S. Effect of combined exposure of diethylphthalate and nonylphenolon on growth of Dunaliella salina. Ecol. Sci. 2012, 31, 370–376. [Google Scholar]
- Wu, W.; Qu, J.; Chen, J.; Hu, G. Toxic effects of nonylphenol ethoxylates and its degradation product on aquatic organisms. J Zhanjiang Ocean. Univ./Zhanjiang Haiyang Daxue Xuebao 2003, 23, 39–44. [Google Scholar]
- Hense, B.A.; Jüttner, I.; Welzl, G.; Severin, G.F.; Pfister, G.; Behechti, A.; Schramm, K.W. Effects of 4-nonylphenol on phytoplankton and periphyton in aquatic microcosms. Environ Toxicol Chem 2003, 22, 2727. [Google Scholar] [CrossRef] [Green Version]
- Guan, C.; SUN, Z.; Min, A.; Duan, S. The ecological toxic effects of Nonylphenol on Phaeocystis globosa. Ecol. Environ. Sci. 2011, 640–645. [Google Scholar]
- Liu, X.; Zhao, J.; Dan, L.X.; Shi, X.; Liang, S. Toxic effects of nonylphenol on dominant microalgae species in Jiaozhou Bay. Mar. Environ. Sci. 2012, 31, 667–673. [Google Scholar]
- Liu, Y.; Dai, X.; Wei, J. Toxicity of the xenoestrogen nonylphenol and its biodegradation by the alga Cyclotella caspia. J. Environ. Sci. 2013, 25, 1662–1671. [Google Scholar] [CrossRef]
- Liu, Y.; Luan, T.-G.; Lu, N.-N.; Lan, C.-Y. Toxicity of Fluoranthene and Its Biodegradation by Cyclotella caspia Alga. J. Integr. Plant. Biol. 2006, 48, 169–180. [Google Scholar] [CrossRef]
- Gao, Q.T.; Wong, Y.S.; Tam, N.F.Y. Antioxidant responses of different microalgal species to nonylphenol-induced oxidative stress. J. Appl. Phycol. 2017, 29, 1317–1329. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Bartosz, G. Oxidative stress in plants. Acta. Physiol. Plant. 1997, 19, 47–64. [Google Scholar] [CrossRef]
- Šepič, E.; Bricelj, M.; Leskovšek, H. Toxicity of fluoranthene and its biodegradation metabolites to aquatic organisms. Chemosphere 2003, 52, 1125–1133. [Google Scholar] [CrossRef]
- Choo, K.; Snoeijs, P.; Pedersén, M. Oxidative stress tolerance in the filamentous green algae Cladophora glomerata and Enteromorpha ahlneriana. J. Exp. Mar. Biol. Ecol. 2004, 298, 111–123. [Google Scholar] [CrossRef]
- Zhou, G.; Peng, F.; Zhang, L.; Ying, G. Biosorption of zinc and copper from aqueous solutions by two freshwater green microalgae Chlorella pyrenoidosa and Scenedesmus obliquus. Environ. Sci. Pollut. Res. 2012, 19, 2918–2929. [Google Scholar] [CrossRef]
- Yan, H.; Ye, C.; Yin, C. Kinetics of phthalate ester biodegradation by Chlorella pyrenoidosa. Environ. Toxicol. Chem. 1995, 14, 931–938. [Google Scholar] [CrossRef]
- Vazquez-Duhalt, R.; Marquez-Rocha, F.; Ponce Rivas, E.; Licea, A.; Viana, M. Teresa Nonylphenol, an integrated vision of a pollutant. Appl. Ecol. Environ. Res. 2005, 4, 1–25. [Google Scholar] [CrossRef]
- Gross-Sorokin, M.Y.; Grist, E.P.M.; Cooke, M.; Crane, M. Uptake and Depuration of 4-Nonylphenol by the Benthic Invertebrate Gammarus pulex: How Important Is Feeding Rate? Environ. Sci. Technol. 2003, 37, 2236–2241. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, N.; Teramoto, T.; Kasai, F.; Sano, T.; Tamaoki, M.; Aono, M.; Kubo, A.; Kamada, H.; Azumi, Y.; Saji, H. Glycosylation of bisphenol A by freshwater microalgae. Chemosphere 2007, 69, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Petroutsos, D.; Wang, J.; Katapodis, P.; Kekos, D.; Sommerfeld, M.; Hu, Q. Toxicity and metabolism of p-chlorophenol in the marine microalga Tetraselmis marina. Aquat. Toxicol. 2007, 85, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.; Soboń, A.; Litwin, A.; Różalska, S. 4-n-nonylphenol degradation by the genus Metarhizium with cytochrome P450 involvement. Chemosphere 2019, 220, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Kang, Y.; Liang, S.; Chen, D.; Zhang, Q.; Zeng, L.; Luo, J.; Jiang, F. Synergistic effect of co-exposure to cadmium (II) and 4-n-nonylphenol on growth inhibition and oxidative stress of Chlorella sorokiniana. Ecotoxicol. Environ. Saf. 2018, 154, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Su, J.Q.; Yang, X.; Zheng, T.; Hong, H. An efficient method to obtain axenic cultures of Alexandrium tamarense—A PSP-producing dinoflagellate. J. Microbiol. Meth. 2007, 69, 425–430. [Google Scholar] [CrossRef]
- Rezaee, M.; Assadi, Y.; Milani Hosseini, M.-R.; Aghaee, E.; Ahmadi, F.; Berijani, S. Determination of organic compounds in water using dispersive liquid–liquid microextraction. J. Chromatogr. A 2006, 1116, 1–9. [Google Scholar] [CrossRef]
- Zhou, G.J.; Peng, F.Q.; Yang, B.; Ying, G.G. Cellular responses and bioremoval of nonylphenol and octylphenol in the freshwater green microalga Scenedesmus obliquus. Ecotoxicol. Environ. Saf. 2013, 87, 10–16. [Google Scholar] [CrossRef]
- Chi, J.; Li, Y.; Gao, J. Interaction between three marine microalgae and two phthalate acid esters. Ecotoxicol. Environ. Saf. 2019, 170, 407–411. [Google Scholar] [CrossRef]
- Sharma, D.K.; Fernández, J.O.; Rosenqvist, E.; Ottosen, C.O.; Andersen, S.B. Genotypic response of detached leaves versus intact plants for chlorophyll fluorescence parameters under high temperature stress in wheat. J. plant. physiol. 2014, 171, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Salin, M.L.; Day, E.D.; Crapo, J.D. Isolation and characterization of a manganese-containing superoxide dismutase from rat liver. Arch. Biochem. Biophys. 1978, 187, 223–228. [Google Scholar] [CrossRef]
- Correa-Reyes, G.; Viana, M.T.; Marquez-Rocha, F.J.; Licea, A.F.; Ponce, E.; Vazquez-Duhalt, R. Nonylphenol algal bioaccumulation and its effect through the trophic chain. Chemosphere 2007, 68, 662–670. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microalgal Species | Regression Equation | R2 | EC50 (mg L−1) |
|---|---|---|---|
| Chlorella vulgaris | y = 0.875x + 0.1257 | 0.975 | 1.534 |
| Scenedesmusobliquus | y = 1.716x + 0.2182 | 0.904 | 1.179 |
| Selenastrumbibraianum | y = 1.6141x + 0.237 | 0.961 | 1.177 |
| Ankistrodesmusacicularis | y = 1.3231x + 0.374 | 0.911 | 1.100 |
| Scenedesmusquadricauda | y = 1.3366x + 0.3974 | 0.961 | 1.080 |
| Chroococcusminutus | y = 1.1814x + 0.4941 | 0.970 | 1.005 |
| Microalgal Species | EC50 (mg L−1) | Reference |
|---|---|---|
| Microcystisaeruginosa | 0.67–2.96 | [27] |
| Dunaliellasalina | 1.47 | [28] |
| Scenedesmusobliquus | 1.0 | [29] |
| Scenedesmussubspicatus | 0.87–0.98 | [30] |
| Phaeocystisglobosa | 0.42 | [31] |
| Skeletonemacostatum | 0.13 | [32] |
| Chaetoceroscurvisetus | 0.22 | [32] |
| Cyclotellacaspia | 0.18 | [33] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, N.; Liu, Z.; Sun, X.; Wang, S.; Liu, W.; Sun, D.; Duan, S. Phytotoxicity, Bioaccumulation, and Degradation of Nonylphenol in Different Microalgal Species without Bacterial Influences. Int. J. Mol. Sci. 2020, 21, 1338. https://doi.org/10.3390/ijms21041338
He N, Liu Z, Sun X, Wang S, Liu W, Sun D, Duan S. Phytotoxicity, Bioaccumulation, and Degradation of Nonylphenol in Different Microalgal Species without Bacterial Influences. International Journal of Molecular Sciences. 2020; 21(4):1338. https://doi.org/10.3390/ijms21041338
Chicago/Turabian StyleHe, Ning, Zhiwei Liu, Xian Sun, Shuangyao Wang, Weijie Liu, Dong Sun, and Shunshan Duan. 2020. "Phytotoxicity, Bioaccumulation, and Degradation of Nonylphenol in Different Microalgal Species without Bacterial Influences" International Journal of Molecular Sciences 21, no. 4: 1338. https://doi.org/10.3390/ijms21041338