Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants



Department of Biological and Environmental Sciences and Technologies, University of Salento, Via Prov.le Lecce-Monteroni, 73100 Lecce, Italy
*
Author to whom correspondence should be addressed.
Insects 2019, 10(12), 441; https://doi.org/10.3390/insects10120441
Submission received: 13 November 2019
/
Revised: 2 December 2019
/
Accepted: 5 December 2019
/
Published: 8 December 2019
(This article belongs to the Special Issue Semiochemicals and Insect Behavior)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:A peculiar feature of all living beings is their capability to communicate. With the discovery of the quorum sensing phenomenon in bioluminescent bacteria in the late 1960s, it became clear that intraspecies and interspecies communications and social behaviors also occur in simple microorganisms such as bacteria. However, at that time, it was difficult to imagine how such small organisms—invisible to the naked eye—could influence the behavior and wellbeing of the larger, more complex and visible organisms they colonize. Now that we know this information, the challenge is to identify the myriad of bacterial chemical signals and communication networks that regulate the life of what can be defined, in a whole, as a meta-organism. In this review, we described the transkingdom crosstalk between bacteria, insects, and plants from an ecological perspective, providing some paradigmatic examples. Second, we reviewed what is known about the genetic and biochemical bases of the bacterial chemical communication with other organisms and how explore the semiochemical potential of a bacterium can be explored. Finally, we illustrated how bacterial semiochemicals managing the transkingdom communication may be exploited from a biotechnological point of view.
1. Introduction
The life of every living being takes place in a dynamic network of relationships with other organisms.
Among the interspecific interactions, those occurring between micro- and macro-organisms have attracted increasing scientific interest and public attention, thus unveiling new and unexpected roles of bugs as beneficial modulators of many biological processes in plant and animal hosts, extending and enhancing their adaptive capabilities.
It is understood that rhizosphere symbiotic microorganisms exert fundamental nutritional and protective effects on different species of plants, strongly contributing to their ecological success [1]. Mycorrhizal fungi provide plants with water and mineral nutrients collected in the soil and counteract soilborne pathogens [2]. N2-fixing bacteria fulfill the nitrogen requirement of several plants in nitrogen-depleted soils [3]. Plant growth-promoting rhizobacteria (PGPR) stimulate plant growth by synthesizing phytohormones and vitamins and exert protective effects on plants against both abiotic and biotic stresses through a variety of well-documented mechanisms [4]. PGPR counteract the water stress by producing exopolysaccharides that alter the soil texture and allow more effective root penetration in the soil. PGPR also sequester and/or degrade toxic compounds, such as heavy metals or xenobiotic aromatic compounds. Moreover, they limit the growth of pathogenic microorganisms and/or protect the plants from their aggression using a variety of mechanisms, including iron limitation by siderophores, the production of antibiotics and extracellular enzymes that attack the microbial cell wall, and elicitation of the induced systemic resistance (ISR) [5]. In turn, in all cases, the host plants provide the symbiotic microorganisms with a protected ecological niche and rhizospheric organic carbon [6]. In insects, symbiotic microorganisms, mostly bacteria, modulate many processes including nutrition, development, immune system, and social and sexual behaviors, making the conquest of new ecological niches possible and, ultimately, promoting the insect evolution [7].
Interestingly, in terrestrial ecosystems, symbiotic microorganisms oversee plant–insect interactions, sometimes according to rather conserved patterns. In general, PGPR exert negative effects on the abundance and activity of herbivore insects, likely due to the priming of primary defenses in the plant [8,9]. In contrast, arbuscular mycorrhizal fungi (AMF) have positive effects on sap-sucking herbivore insects. This is likely due to positive effects of symbiotic fungi on the nutrient level in the phloem [8,9]. AMF effects on chewing herbivore insects are, instead, generally neutral. However, when present, the effects are beneficial for specialist chewing herbivores and detrimental for those of the general kind. Furthermore, AMF generally exert negative effects on natural plant enemies [10]. In the establishment of specialized plant–insect interactions, such as in the case of the interaction between Mentha aquatica and Chrysolina herbacea [11], an aspect of great interest is represented by the dynamics of plant- and insect-associated microbial communities. This also applies to the role of bacterial volatile organic compounds (BVOCs) in regulating the crosstalk between plants and generalist herbivores and in protecting plants from generalist herbivores.
Indeed, an essential aspect of the interspecific interaction is that of communication that triggers and controls the specific biological responses that underlie the symbiosis. Recently, scientific literature has attributed an increasingly important role to the microbial volatile organic compounds (MVOCs) and, in particular, of BVOCs in the chemical communication between microorganisms, plants, and insects [12,13]. There are a number of examples illustrating how secondary metabolites produced by symbiotic bacteria may act as intraspecific, interspecific, and mating signals in insects affecting their sexual and social behavior, and how bacterial secondary metabolism and plant secondary metabolism may be functionally interconnected between them to produce chemical signals that may be active toward specific insect populations [14,15].
Bacteria play surprising roles in presocial and eusocial insect intraspecies communication. Presocial skills, which are defined as simple phenomena mediated by pheromones, are very diffuse in many insect orders. Specific BVOCs, which are mostly produced by gut bacteria, work as multifunctional pheromones in presocial insect communication. More sophisticated mechanisms underlie eusocial insect communication, which supports reproduction, the development from egg to adult, and influences the behaviors during all life stages. Hymenoptera, particularly ants, bees, and wasps, are the most well-known examples of eusocial insects. These insects developed complex social skills during evolution, implementing a complex network between individuals and forming nests. Cuticular hydrocarbons (CHCs) play a key role in the social insect communication, and there is some evidence that the social insect microbiota may be involved in CHC metabolism [16]. As well as in presocial insect communication, MVOCs, in particular BVOCs produced by gut bacteria, are involved in eusocial insect and sexual communication. Also, positive (mutualism) or negative (parasitism) interspecies relationships are established using mechanisms and compounds (BVOCs) related to social communication in insects [17].
Elucidating the biological role, the biochemistry and regulation of the chemical signals produced by microorganisms is not only essential to better understanding the ecological significance of transkingdom interactions involving microorganisms, plants, and insects, but it is also instrumental to the development of biotechnological devices. This tool is imperative for understanding the biological struggle for more sustainable agriculture and to deal with the effects of the climate changes on plant–insect ecosystems, which inevitably involve changes in microbial community dynamics with a far greater knowledge [18].
2. Interactions and Chemical Crosstalk between Microorganisms, Plants, and Insects: Ecological Models and Study Cases
Transkingdom interactions involving microorganisms, plants, and insects are generally complex, multifaceted, and arduous to analyze. They involve multiple organisms at different macro- and micro-scale levels, thus, taking transkingdom interactions beyond the classic schemes exemplified by the major types of biological interactions involving pairs of organisms and often causing them to be conditional and affected by abiotic environmental parameters [19]. Consequently, the chemical crosstalk that oversees transkingdom interactions is sophisticated and hard to decipher from an ecological point of view. Another concern when studying the role of microbial semiochemicals in transkingdom interactions is relating the chemical signals that a microorganism can produce at a micro-scale level in a natural environment to a biological response in a macro-organism [20]. Therefore, the setup of appropriate ecological models to consolidate results is strongly encouraged. Below, we describe several ecological models suitable for studying the interactions between microorganisms, plants, and insects, along with the established or hypothesized role of compounds of microbial origin in the transkingdom communication. Here, the role of bacteria and products of their metabolism in intraspecies presocial and eusocial insect communication, in sexual communication, and in interspecies communication supporting mutualism or parasitism is illustrated.
2.1. Bacteria and Presocial Insect Communication
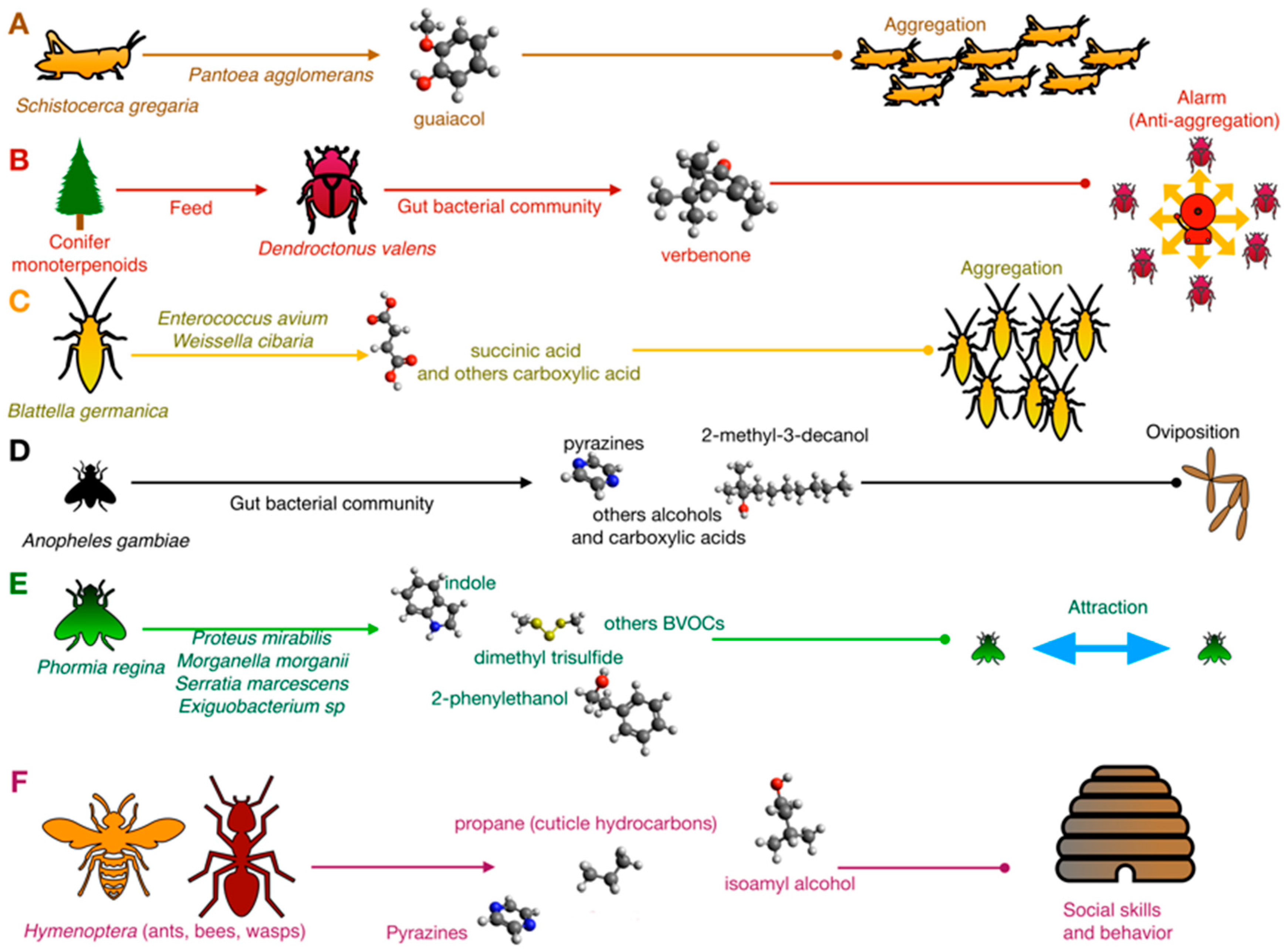
The bacterial contribution to chemical crosstalk was first investigated in simple phenomena such as attraction/cohesion processes or sexual behavior. For example, Dillon and colleagues [21,22] demonstrated that in the desert locust Schistocerca gregaria (Orthoptera, Acrididae), the aggregation pheromone guaiacol was produced in vitro by Pantoea aggolomerans isolated from the insect gut using vanillic acid introduced by diet as a substrate (Figure 1).
Microbial biotransformations are, indeed, rather common in insects. In the bark beetle Dendroctonus valens (Coleoptera, Curculionidae), the most important mortality agent in the coniferous forest, the gut microbiota transforms toxic monoterpenoids of conifers into the multifunctional pheromone verbenone [23,24]. In D. valens, the verbenone works as an aggregation pheromone at a low concentration, while it is an alarm (anti-aggregation) pheromone at a high concentration. Thus, it has been proposed for coniferous forest management. Gut bacteria transform cis-verbenol, a toxic precursor for bacteria, to verbenone. It subsequently benefits from this conversion due to the reduced toxicity of verbenone. At the same time, by producing and modulating the bark beetle ambivalent pheromone, they play a key role in controlling D. valens population density (Figure 1). Specific groups of terpene-metabolizing bacteria that are associated with monoterpene tolerance were described in different Dendroctonus species. In D. valens, the terpene-metabolizing bacteria were almost equally represented by both Gram-positive (Actinobacteria, Bacilli) and Gram-negative (Gamma-Proteobacteria) species, while in D. ponderosae, the Gram-positive species were predominant. Compared to the bacteria isolated from D. valens, the bacteria from D. ponderosae were not able to grow using α-pinene and 3-carene as carbon substrate. They were therefore inhibited by different monoterpenes [24], suggesting that a reduction in Gammaproteobacteria may affect their tolerance to monoterpenes. A different study reported that several Serratia strains from D. ponderosae were able to degrade monoterpenes but not α-pinene that was, instead, degraded by Rahnella. On the other hand, Serratia and Brevundimonas were able to reduce diterpenes [25]. Compounds and strains involved in these systems are reported in Table S1 of Supplementary Materials.
The upmost importance of bacteria in insect aggregation was also demonstrated in the German cockroach Blattella germanica (Blattodea, Blattellidae). This cockroach is a major pest of the built environment, where it can acquire and transmit pathogens and produce allergens that cause disease in humans. Wada-Katsumata and coworkers [26] demonstrated that the feces of normal cockroaches emitted volatile carboxylic acids (VCAs) that elicited insect aggregation, whereas bacteria-free feces contained few VCAs and were relatively unattractive. Among bacteria, Enterococcus avium and Weissella cibaria were shown to produce aggregation BVOCs 3-isovaleric acid and valeric, succinic acid, benzoic acid, and phenylacetic acid [26] (Figure 1).
A similar mechanism was also demonstrated in the black blow fly Phormia regina (Diptera, Calliphoridae), in which bacteria of the species Proteus mirabilis, Morganella morganii, Serratia marcescens, and Exiguobacterium spp. emit strain-specific BVOCs (Table S1) which work as attraction signals [27] (Figure 1).
Many studies were focused on aphids (Rhynchota, Aphidiae), a common group of plant parasites that live in a colony and feed on plant sap releasing honeydew, a sugar-rich, dense liquid used by Hymenoptera as a feed source. Different semiochemicals dealing with intraspecies communication are present in honeydew, including the E-β-farnesene and a number of BVOCs (Table S1). E-β-farnesene is an alarm pheromone [28,29,30] and is a key component of communication. Sixteen out of 26 aphid species emit this compound as the only or among the most common volatiles [31].
In insects, BVOCs are also signals for oviposition (Figure 1). In Anopheles gambiae (Diptera, Culicidae), an African mosquito considered one of the most effective vectors of malaria, an inoculum of gut bacteria stimulated oviposition through semiochemicals 2-methyl-3-decanol 24, 3-methyl-1-butanol 21, 2-phenylethanol 11, indole 9, 3-methylbutanoic acid 25, diisopropylpyrazine 26, 2,5-diisopropylpyrazine 27, isopropyl-secbutylpyrazine 28, isopropyl-isobutylpyrazine 29, phenylmethanol 30, and 2-phenylethanol 31 [32,33] (Table S1). Other behavioral responses of this mosquito are based on semiochemicals signature produced by the microbiota: BVOCs released by human microbiota on the skin surface drive the choice of the host, influencing the diffusion of malaria directly [34]. Staphylococcus epidermidis releases various BVOCs that influence this phenomenon, such as 3-methyl butanal, 2-methyl butanal and 2-methylbutanoic acid, and 3-methyl-1-butanol and 3-methylbutanoic acid [35]. A similar phenomenon occurs in yellow fever vector Aedesa egypti, in which the relationship between the attraction of adult female individuals and quorum sensing regulation in S. epidermidis was investigated [36].
2.2. Bacteria and Eusocial Insect Communication
In social nest-living insects, BVOCs regulate social behaviors. For example, in Atta sexdens (Hymenoptera, Formicidae), 2,5-dimethylpyrazine and 3-ethyl-2,5-dimethylpyrazine were identified in poison gland secretion and work as powerful trail pheromones (Figure 1). In A. sexdens var. rubropilosa (Hymenoptera, Formicidae), Serratia marcences, a common endosymbiotic strain in insects, is involved in pyrazines biosynthesis starting from L-threonine as a precursor [37]. In addition to bacteria, yeast species also contribute to semiochemical communication in Hymenoptera. In North American yellowjacket wasps (Hymenoptera, Vespidae), two species of fungi produce VOCs that attract conspecific individuals. Hanseniaspora uvarum and Lachancea thermotolerans were attractive to three species of yellowjacket wasps (V. pensylvanica, V. germanica, and V. alascensis) when grown on grape juice-infused yeast peptone dextrose (YPD) agar [38]. Gas chromatography–mass spectrometry analysis revealed that ethyl acetate (only H. uvarum), phenylethyl acetate, isoamyl acetate, 2-phenylethyl alcohol, isoamyl alcohol, and 2-methyl-butanol were the most abundant headspace volatiles produced by these yeasts. This information was instrumental to formulate a synthetic blend that was proven to be effective in attracting V. pensylvanica, but not the other yellowjacket species [38].
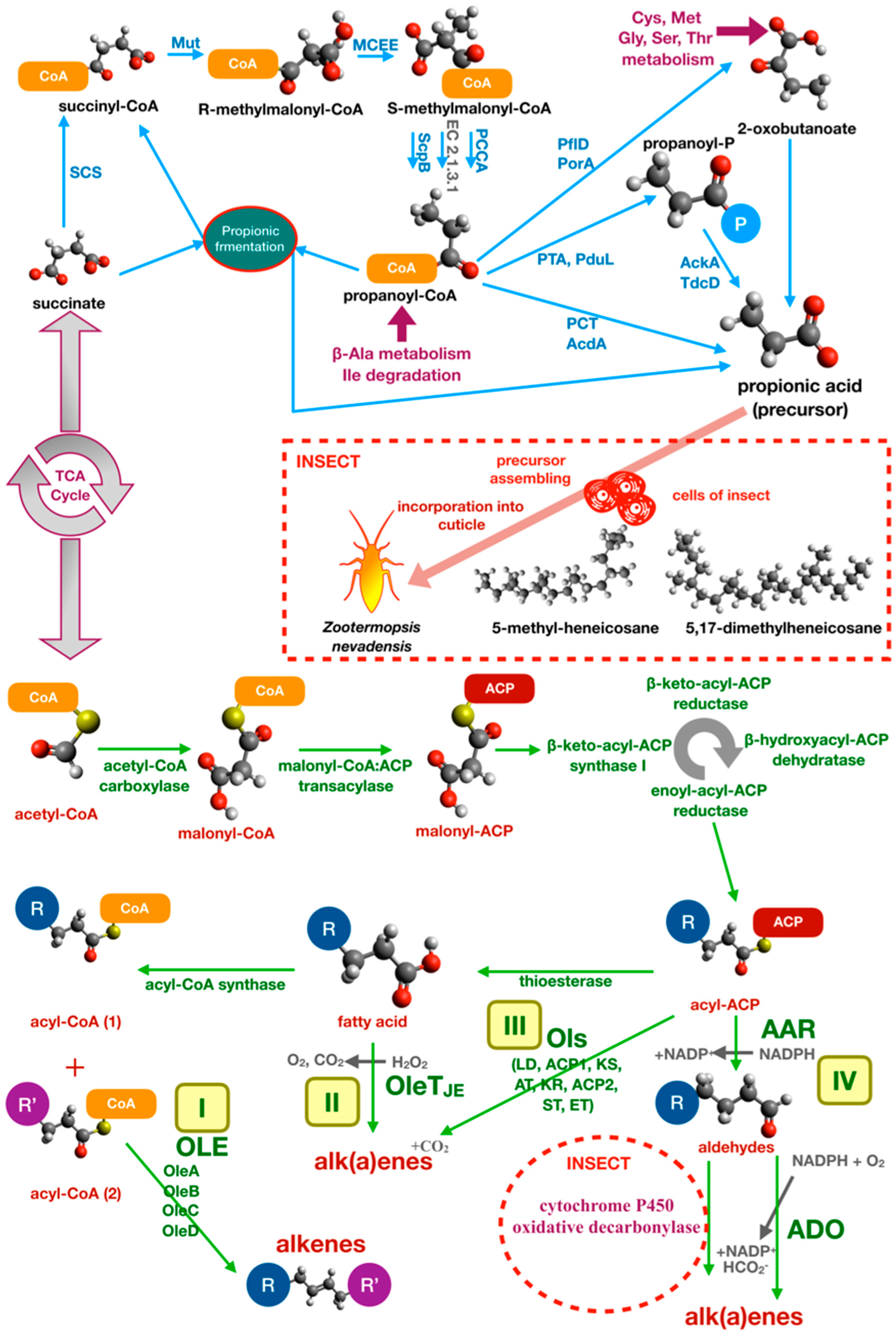
Another important type of communication in social insects is based on chemical recognition of CHCs. In termite species, the aggressive response is common when a parasite or non-nestmate insect penetrates the nest. In Reticulitermes speratus (Isoptera, Rhinotermitidae), antibiotic feeding increases the mortality and aggression of test individuals [39]. Similar effects were obtained with the addition of extracts from gut bacteria of non-nestmate individuals [39]. The death or reduction of microbial symbionts also affects the fitness of individuals, showing a reduction in body size and in mechanical resistance of the exoskeleton formed by a thinner cuticle [40]. Direct evidence of CHC metabolism by gut bacteria was obtained in Zootermopsis nevadensis (Isoptera, Termopsidae) using radiolabeled succinate, a precursor of methyl-branched hydrocarbons, which was transformed by bacteria into propionate and incorporated in the cuticle as 5-methyl-heneicosane or 5,17-dimethylheneicosane by insect cells [41]. A specific analysis of the effects of gut microbiota on cuticular hydrocarbon metabolism in ants was performed by Teseo and colleagues [41] based on three point-of-view approaches. These experiments investigated the relationship between variation in bacterial community, differences in CHC profiling, and nestmate recognition. The authors manipulated the gut microbiota of Acromyrmex echinatior leaf-cutting ants (Hymenoptera, Formicidae) using tetracycline. Before partially restoring the microbiota through fecal droplet administration, they analyzed the relationship between microbiota structure and insect survival and found that survival was enhanced by endosymbionts. Moreover, they proved that antibiotic treatment modified the normal microbiota structure and affected the CHC chemical profile. Additionally, results showed that modified CHC chemical profile was significantly correlated with enhanced aggression and misrecognition. Variations in CHC levels were significant for 4-oxo-octanoic acid, 4-oxo-decanoic acid and in long-chain linear alkanes (n-C36 and n-C40), which decreased in antibiotic-treated insects compared to untreated controls [42].
Finally, in eusocial insects, the gut microbiota assemblage reflects the exposure to social interactions: Oral trophallaxis enhanced the development a healthy “core microbiota” in the bee. Gram negative bacteria, such as Gilliamella apicola, Frischella perrara, and Snodgrassella alvi, are acquired with either social interaction between young individuals, nurse/workers individuals, or those that have contact on hive surfaces. These studies have shown how the social contact may affect the normal gut microbiota in the virgin queen bee [43,44]. Other studies demonstrated that the Apis microbiota is caste-specific [45,46] and age-specific [47]. It is also affected by the environmental landscape [48]. Queens and workers harbor strikingly different gut microbiota, and the queen replacement affects the microbiota in workers [47]. These phenomena can explain that any nest or group of insects has a specific volatile profile, a specific microbiota, and a specific cuticular profile that acts as molecular signature and contributes not only at the intraspecific recognition, but also as social and behavior skills at intranest level.
2.3. Bacteria and Sexual Communication in Insects
Sexual recognition in the insect is another intriguing phenomenon influenced by bacteria. Specifically, the microbiota can influence reproduction with different mechanisms, such as sexual manipulation or pheromone and semiochemical manipulation. Sexual manipulators include bacteria of the genera Wolbachia, Rickettsia, Cardinium, Spiroplasma, and Arsenophonus. They affect reproduction in different ways: They may induce parthenogenesis, feminization, or male killing [49]. On the other hand, a number of bacteria and some viruses have the ability to influence the sexual behaviors.
In an interesting model, the Hz-2v virus was shown to enhance sexual pheromone production in Helicoverpa zea (Lepidoptera, Noctuidae) females that attracted more males than non-infected females, thereby facilitating virus transmission [50]. Meanwhile, the bacterium Staphylococcus aureus was shown to manipulate pheromone levels in male boll weevils thanks to the activity of Enterotoxin B [51].
In Costelytra zealandica (Coleoptera, Scarabaeidae), symbiont bacteria belonging to the species Morganella morganii isolated from colleterial glands produce phenol form tyrosine as a female sexual pheromone [52,53]. The CHC chemical profile can also regulate sexual interactions between ants: Male individuals of Cardiocondyla obscurior (Hymenoptera, Formicidae) genus mimic CHCs of females in sexual conflict [54,55], and an altered gut microbial community causes leaking of tyrosine precursor 4-hydroxyphenylpyruvate for cuticle formation. This precursor is provided by the Westeberhardia bacterial symbiont [56].
In the plant–insect systems formed by Chrysolina herbacea (Coleoptera, Chrysomelidae, Chrysomelinae) and Mentha aquatica, diet and sexual identity are key factors that drive sexual recognition between individuals [57]. M. aquatica adopt a constitutive defense system against herbivore based on terpenoids biosynthesis in glandular trichomes [58]. Some terpenoids regulate the attraction of insects: Pulegone, a major compound emitted by undamaged plants, is a powerful attractant for C. herbacea, while menthofuran works as a repelling agent produced by damaged plants [11]. GC–MS analysis of M. aquatica oils before and after the inoculum of bacterial consortia isolated from the insect gut revealed as BVOCs amount and type are linked to the sex of individuals [57]. In the microbial community from female individuals 3-nonanol, β-bourbonene, isopulegol, α-amorphene, and longipinanol were detected, while 3-nonanol and isopulegol were detected in the microbial community from males. Both communities could biotransform the essential oil by producing menthofuran, menthone, 1,8-cineole, limonene, and pulegone. Several of these compounds were previously described as pheromones in insects. For instance, in Crematogaster sjostedti (Hymenoptera, famiglia), 3-nonanol mediates alarm, defense response, and conspecific recognition [59]. In Azteca chartifex (Hymenoptera, famiglia), α-amorphene is detected in pygidial gland secretions, while β-bourbonene is a pheromone in Euceraphis punctipennis (Rhynchota, Aphidiae) [31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60].
2.4. Bacteria and Interspecies Insect Communication
As described before, microorganisms may influence chemical signaling with BVOC emission or CHC metabolism in insect communities. In a number of mutualistic symbioses, as well as in some parasitic interactions between insect species, the acquisition and exchange of microbial strains can induce a phenomenon of chemical camouflage that reduces the aggressiveness between non-nestamate insects [61].
In literature, many reported observations of mutualist relationships were focused on ants for different reasons: (I) Simple feeding and growth, and easy management under laboratory conditions; (II) direct impact on agriculture by increasing soil quality and nutrients; (III) indirect impact in biocontrol of pest insects. The most well-known example of microbial-mediated mutualism in ants is that which is established between ants and aphids (Rhynchota, Aphidiae). Ants establish a long-term mutualistic interaction with aphids, managing the aphid nest and protecting them from natural aphid predators and parasites. In turn, by feeding on plant sap, aphids produce honeydew for the ants [62].
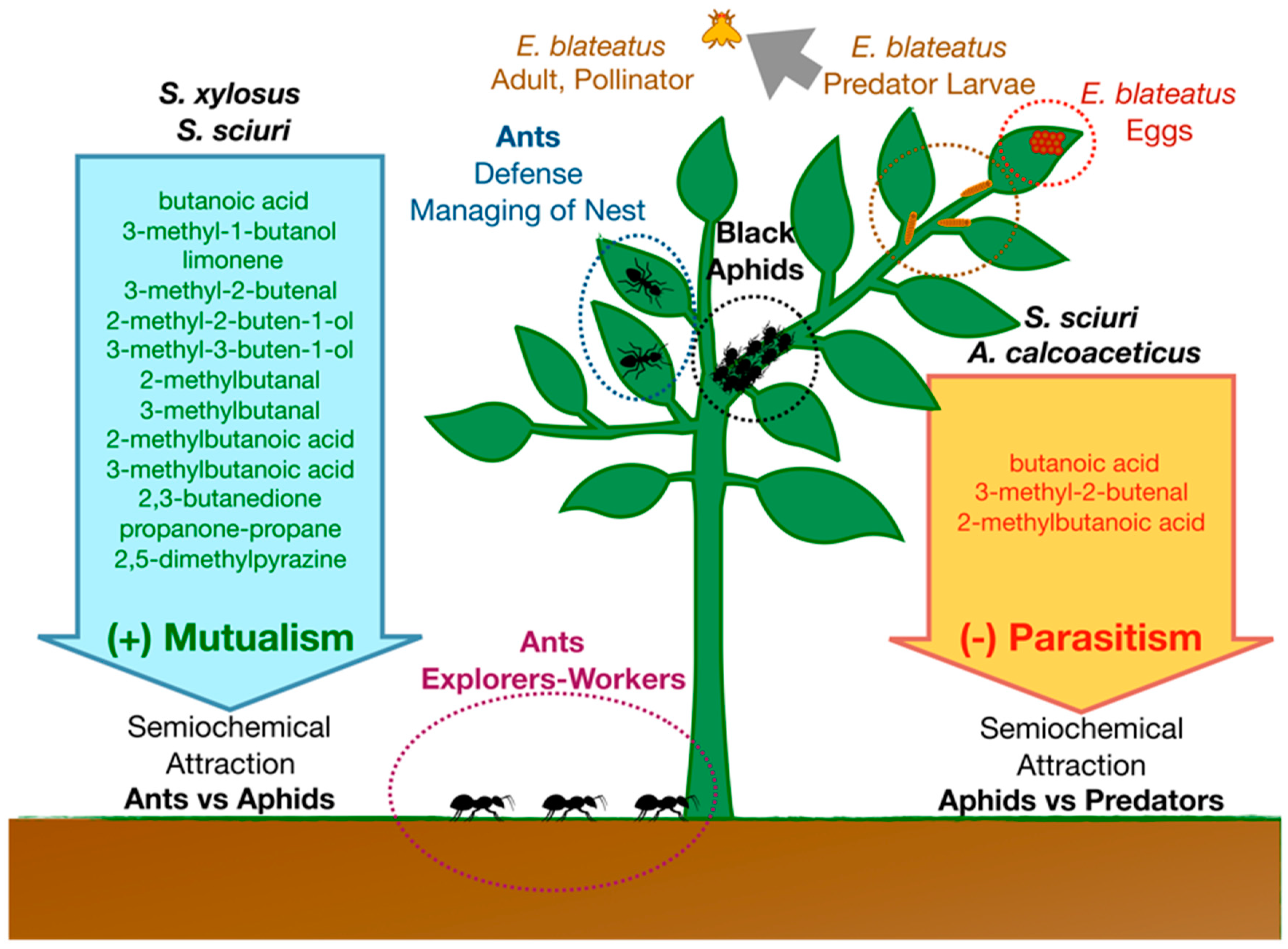
In a well-characterized ant–aphid interaction model, BVOCs produced by Acyrthosiphon pisum (Rhynchota, Aphidiae)-associated Staphylococcus sciuri and Staphylococcus xylosus living in the honeydew produced by aphid mediate mutualistic interaction with Lasius niger (Hymenoptera, Formicidae) [29,30]. Bacteria produce a blend of semiochemicals (including limonene, butanoic acid, 3-methyl-1-butanol, 3-methyl-2-butenal, 2-methyl-2-buten-1-ol, 3-methyl-3-buten-1-ol, 2-methylbutanal, 3-methylbutanal, 2-methylbutanoic acid, 3-methylbutanoic acid, 2,3-butanedione, propanone-propane, 2,5-dimethylpyrazine) that attract ant scouts (Figure 2). However, at the same time, some semiochemicals produced in this system by S. sciuri and Acinetobacter calcoaceticus (butanoic acid, 3-methyl-2-butenal, 2-methylbutanoic acid) act as attractants and ovipositional stimulants for the natural enemy of A. pisum, Episyrphus balteatus (Diptera, Syrphidae). This is one of the most efficient, abundant, and highly specialized aphidophagous predators [63].
Another type of insect–insect mutualistic interaction is that between ants and butterflies during their larval stages. Interactions can be facultative or obligate, mutualistic or parasitic. In the interaction between Myrmica (Hymenoptera, Formicidae) and Maculinea (=Phengaris) (Lepidoptera, Lycaenidae), ants defend instars from natural enemies (usually staying on top of instars). In turn, they are rewarded with nutrition secretions from specialized exocrine glands [64]. On the other hand, this relationship can be parasitic. Caterpillars of some Maculinea species are adopted by certain Myrmica species, transported into their nest, and fed on either ant regurgitations (trophallaxis) or directly on ant brood [65]. The central role of CHCs and acoustic signals in the crosstalk between butterflies and ants is well-known [66]. The possibility that symbiotic microorganisms living on butterflies and ants may influence this host–parasite interaction by either CHC metabolism or MVOC production has recently brought the study of the butterfly and ant microbiota to attention [67,68].
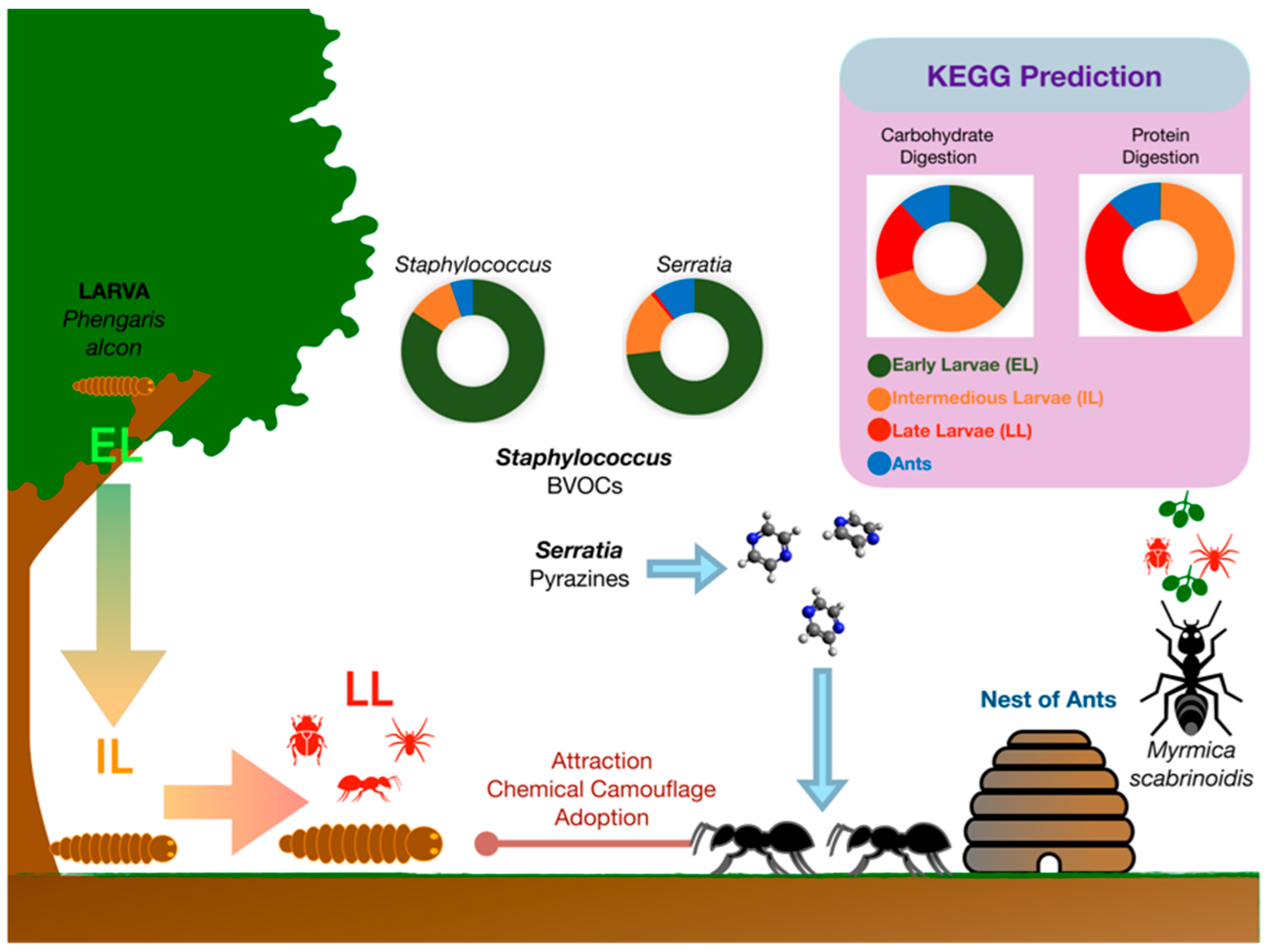
The gut microbiomes of the endangered butterfly Maculinea alcon at different life stages and that of its adopting ant Myrmica scabrinodis were characterized in a recent study case [67]. On the Alpine Arc, the life cycle of Maculinea alcon begins in August when eggs are deposited on both flower buds and leaves of Gentiana asclepiadea or Gentiana pneumonanthe. Early larves (EL) hatches through the base of the egg into the flower ovary, where they spend two to three weeks eating the flower tissue and developing seed. Phytophagous EL moults three times. Then, they chew a hole in the flower, through which they let themselves fall into the soil. Here, intermediate larvae (IL) wait until they are adopted by the ants that collect the IL and carry them to their nest. Late larvae (LL) are fed by worker ants and become carnivorous. They spend the whole autumn, winter, and spring in the ant nest. Then, in the early summer, they turn into pupae. A month later, adult butterflies emerge from pupae, which leave the ant nest and fly to flowers in July for a one-month duration [69] (Figure 3).
As demonstrated by 16S rRNA-guided metabarcoding, the M. alcon larval development is associated with significant changes in the structure and predicted functions of the butterfly microbiota [67]. Alphaproteobacteria dominated M. alcon microbiota during EL and IL stages, while gamma-proteobacteria were predominant in LL. The PICRUSt functional prediction from DNA metabarcoding data suggested changes in predominant biochemical pathways during larval development, which are consistent with a different ability to digest carbohydrates (herbivorous EL) or proteins (carnivorous LL) and to metabolize CHCs (Figure 3). A characteristic pattern was represented by an increase, among the intracellular obligate bacterial endosymbionts, in Rickettsiella (Gammaproteobacteria) abundance during larval development and a parallel decrease in Wolbachia (Alphaproteobacteria) abundance. This shift may be associated with larval color shift from red (EL) to green/yellow (LL). Indeed, Rickettsiella is known to be responsible for a red to green color shift in pea aphids, which makes the aphids less visible to predators and parasites [70]. A similar red to green/yellow color shift may facilitate the integration of M. alcon larvae in the host ant colonies considering that ant species have a dichromatic color vision system that is insensitive to red light [71].
Another characteristic pattern was represented by dynamics of BVOC-producing Serratia spp., Staphylococcus spp., and actinomycetes in M. alcon larvae. In particular, Serratia marcescens and Serratia entomophila were largely abundant in EL, while their abundance progressively declined in IL and LL. Microorganisms belonging to the genus Serratia are very common in plants, where they act as plant growth-promoting bacteria, as well as in the gut of arthropods as symbiotic bacteria. Intriguingly, these bacteria produce volatile pyrazines used as trail pheromones by insects. Particularly, 2,5-dimethylpyrazine and 3-ethyl-2,5-dimethylpyrazine are trailmarkers in ants. Thus, the abundance of S. marcescens and S. entomophila in EL and IL may play a specific role in the production of deceptive pyrazines that facilitate recognition of the M. alcon larvae by adopting ants.
2.5. Bacteria and Insect–Plant Communication
As exemplified above with the Chrysolina herbacea—Mentha aquatica model, bacteria play a pivotal role in plant–insect interaction systems. The vetiver grass (Vetiveria zizanoides (L.) Nash, syn. Chrysopogon zizanoides (L.) Roberty) provide us with another intriguing example. Vetiver is the only grass cultivated worldwide for to its multiple properties. The vetiver grass is very suitable for soil erosion management [72], sustaining agricultural productivity [73], and phytoremediation [74,75]. From the long roots of vetiver, an essential oil is extracted, which is used to formulate many perfumes. In addition, vetiver is extremely resistant to (or sometimes repellent to) many insect pests, although it may be often infested by pest of other crops, such as stem borers, white grubs, cicadas, and termites, which use vetiver as a refuge [76,77,78]. A study focused on Chilo partellus, a lepidopterous stem borer of grasses that may infest the vetiver and is a serious pest to maize, sorghum, rice, and other crops, showed that the vetiver grass is highly preferred for oviposition with respect to the aforementioned crops. However, larval survival on vetiver was extremely low, raising the possibility that this plant can be used as a plant trap around crops on which stem borers are a problem [79,80]. There is evidence suggesting that many insects use vetiver grass as refuge, and that the insect biodiversity of vetiver is actually very high including not only potential insect pests, but also a large number of general predators and parasitoids of insects [81]. The ability of the vetiver grass to attract (and manage life cycle) or repel numerous species of insects relies on the high plant potential for biosynthesis of secondary metabolites. The biosynthesis of these compounds is particularly intense in the vetiver roots that are traditionally used to repel cloth moths, head lice, and bed bugs. The vetiver oil that is extracted from the vetiver roots is a complex mixture of hundreds of sesquiterpene alcohols and hydrocarbon compounds. Several of which possess insect repellent properties. α-vetivone, β-vetivone, khusimone, zizanal, epizizanal, and (C)-(1S, 10R)-1,10-dimethylbicyclo (4,4,0)-dec-6-en- 3-one) were reported to be repellent to different insects. In particular, zizanal and epizizanal have topical irritant activity on cockroaches and flies [82]. Nootkatone, zizanol, and bicyclovetivenol are strong repellents and toxicants to Formosan subterranean termites Coptotermes formosanus (Isoptera, Rhinitermitidae) [83]. Indeed, the efficacy of vetiver oil and nootkatone as soil barriers against these termites was proven [84].
Intriguingly, there is evidence that root-associated bacteria may be involved in the biosynthesis of several constituents of the vetiver oil. These bacteria, living in the essential oil-producing cells and in the lysigen lacunae in close association with the essential oil, metabolize biosynthetic precursors that are synthetized by the plant [85,86]. This is supported by evidence that axenic vetiver produced only trace amounts of oil in vitro, with a strikingly different composition compared to the oils from in vivo vetiver plants. Most root-isolated bacteria were grown using oil sesquiterpenes as a carbon source and were biotrasformed into a large number of compounds typically found in commercial vetiver oils [85] (Table S1). Some of these compounds have repellent activity to insects, such as bicyclovetivenol, which was synthesized in vitro by vetiver root-associated Pseudomonas sp. VET-3 and Pseudomonas sp. VET-5 when fed on plant precursor β-caryophyllene, one of the few compounds produced by axenic vetiver [86]. This finding is consistent with a leading role of vetiver root-associated bacteria in overseeing the complex interaction between the plant and its hosted insects. Other volatile terpenoids produced by vetiver root-associated bacteria are listed in Table S1.
Plant–insect mutualism, based on pollination performed by Hymenoptera, is established thanks to the microbiota of nectar. Rering and colleagues [87] demonstrated that yeasts (Metschnikowia reukaufii and Aureobasidium pullulans) and bacteria (Neokomagataea sp. and Asaia astilbes) produce volatiles compound in nectar. All microbial strains produced n-hexanol, while only bacteria produced the characteristic compound 2,5-dimethylfuran, while alcohols, esters, and ketones were more abundant among fungal metabolites [87]. In plant Hymenoptera systems, bacterial symbionts are also involved in seed preservation. The first insect-produced herbicide myrmicacin, which was isolated from the ants, stopped seed germination and preserved the seeds from structural damages [88]. Myrmicacin analogs play similar roles. These compounds can also stop pollen germination and mitotic division in plants and fungal cells and may inhibit fungal growth on seed surfaces [89]. Some Lactobacillales, such as Lactobacillus plantarum, produce myrmicacin-related hydroxy acids [90]. All these functions can be counteracted by the production of antibiotics by Actinomycetales in the “fungal garden”, a specialized structure managed by ants that cultivate fungi as a source of food [91].
Another interesting point is the ability of nonpathogenic bacteria to mediate physiological response and signaling pathways in plants. A review by Dicke shows that phytobiome, which is formed by microbiome and macrobiome (insect), manages phenotypic plasticity in plants, inducing changes in VOCs emitted by the plant and influencing the relationship between herbivores and plants [92]. Some nonpathogenic bacteria cause transcriptomic and metabolic changes in plants. For example, root-colonizing Pseudomonas fluorescens SS10 induces resistance against another pathogenic Pseudomonas species (P. syringae pv tomato, Pst), but also enhances resistance against the insect pest Spodoptera exigua (Lepidoptera, Noctuidae). This induction of resistance depends on a signaling pathway based on salicylic acid, while other signaling pathway systems are based on phytohormones as jasmonic acid and ethylene. Using Arabidopsis thaliana, Van de Mortel and coworkers identified approximately 50 metabolites related to the salicylic acid signaling pathway and demonstrated that camalexin and glucosinolate were principal compounds related to resistance against Pst, using gene-disruption approach to silence signaling [93].
Another study showed that the root colonization by P. fluorescens in A. thaliana influences not only the defense plant response to herbivores, but also the relationship between herbivores and their parasitoid. As shown in experiments with A. thaliana, the VOCs profile emitted by plants infested by aphid Myzus persicae (Hemiptera, Aphididae) is less attractive for the parasitoid Diaeretiella rapae (Hymenoptera, Braconidae) if the roots of the plant are colonized by root bacteria [94]. This phenomenon is lost when the jasmonic acid production is silenced in the plant [94]. The jasmonic acid signaling pathway is related also to the mediation of plant defenses in bitrophic systems. Caterpillars of Pieris brassicae (Lepidoptera, Pieridae) develop faster and bigger when the plants are simultaneously infected by aphid Brevicoryne brassicae (Hemiptera, Aphididae). This phenomenon can occur because the infestation of aphids reduces the levels of jasmonic acids by ten-fold as a consequence of low transcriptions of the lox and myc genes, coding for enzyme-related to jasmonic acid biosynthesis [95]. The facilitation also affects the way herbivores influence parassitoids. As reported in the example described previously, the development of the parasitic wasps Cotesia glomerata (Hymenoptera, Braconidae) is faster if the parasitized caterpillars are fed on by plant–infested aphids, probably because a better fitness of herbivorouse hosts leads to a better fitness of parasitoid [95].
The third trophic level, represented by parasitoid of herbivores, appears to be most influenced by symbionts. C. glomerata injects in the egg of herbivores, such as P. brassicae and P. xylostella (Lepidoptera, Plutellidae), venom, and the symbiotic polydnaviruses (PDVs) that sustain the development of parasitoid. This interaction caused an altered plant-mediated interaction that reduced colonization of other herbivores [96]. On the other hand, symbiotic interaction can protect the pray to parasitoid as shown for aphids, attenuating herbivore-induced plant volatiles [97,98].
3. BVOC Biosynthetic Pathways and Their Intersections with Primary and Secondary Metabolism
The volatile molecules produced by bacteria play a primary role in the chemical communication between insects. They share common features including low molecular mass, high vapor pressure, low boiling point, and low polarity, although they may be produced through very different biosynthetic pathways. The aim of this section was to connect entomology to industrial microbiology using a biochemical point of view, implementing a classification method for semiochemical compounds based on microbial source and pathway of biosynthesis (Table S1). The mechanisms of biosynthesis are also interesting for an industrial purpose: The fermentation of biomass to produces semiochemical useful to manage pests and insects. In this section, we attempted to illustrate some validated or proposed pathways underlying the production of these compounds. The output was an atlas (Table S1) that links each BVOC that acts as a signaling molecule in insects to the respective producing microorganism(s) and biosynthetic pathway(s). For this purpose, the atlas was hyperlinked to both KEGG and Pherobase databases. Excel’s formatting allowed a suitable sorting of data according to consultation needs. Some biosynthetic pathways led to the production of BVOCs through primary and secondary metabolism. These pathways are illustrated below.
3.1. BVOCs and Primary Metabolism
A number of BOVCs may be produced as intermediates, byproducts, or endproducts of the bacterial primary metabolism through well-conserved pathways. Most of these pathways are fundamental for the energy metabolism, for the biosynthesis of cellular structure, and for bacterial growth and survival. Simple carboxylic acids, such as formic acid, acetic acid, lactic acid, and succinic acid, may be produced by mixed acid fermentation in facultative anaerobic enteric bacteria. When their intracellular concentration is high as a result of an intense metabolic activity, the molecules are released passively in the environment as BVOCs. Other enteric bacteria may vigorously produce acetoin (3-hydroxybutan-2-one) and its reduced form 2,3-butanediol through butanediol fermentation. Several clostridia possess the acetone-butanol-ethanol fermentation that produces propanone and butanol. Clostridia are strictly anaerobic bacteria [99] that, thanks to this peculiar energy metabolism pathway, are used to produce solvent or fuels including hydrogen [100]. Propionibacterium- and propionate-fermenting microorganisms are other bacteria with a peculiar energy metabolism that contributes to propionate production from succinate. Many fermentative pathways lead to acetate production in addition to other metabolic processes such as those occurring in strictly anaerobic homoacetogenic bacteria.
Amino acid metabolism by the Ehrlich pathway [101] through the catabolism of branched-chain amino acids (leucine, valine, and isoleucine), aromatic amino acids (phenylalanine, tyrosine, and trytophan), and the sulfur-containing amino acid (methionine) leads to the formation of fusel acids and fusel alcohols, and many of these BVOCs may act as semiochemicals. In particular, 2-phenylethanol and phenylacetic acid are produced by phenylalanine catabolism. Additionally, guaiacol and phenol are formed through tyrosine catabolism, while the indole is a direct precursor of tryptophan. Many BVOCs can be produced through the catabolism of branched-chain amino acids. It is interesting to note that research in this field intersects with biotechnology. A number of compounds, which act as chemical signals, are also studied for some biotechnological applications. An example is biofuel production using Corynebacterium glutamicum that was engineered for producing 2-methyl-1-butanol and 3-methyl-1-butanol via the Ehrlich pathway from 2-keto-3-methylvalerate and 2-ketoisocaproate, respectively [102]. Another example is provided by an E. coli strain that was genetically engineered to implement the production of 2-methyl-1-butanol, 3-methyl-1-butanol and 2-phenylethanol using 2-keto-isocaproate 2-keto-methyl-valerate 2-keto-isovalerate 2-phenyl-pyruvate as a substrate [103,104].
Volatile sulfur compounds are produced through methionine catabolism. Methionine gamma-lyase is central in this catabolic pathway. This enzyme directly degrades sulfur-containing amino acids to α-cheto acids, ammonia, and thiols [105]. Methionine is decomposed by various bacteria with production of methanethiol, dimethyl sulfide, and dimethyl disulfide [106,107]. All these compounds, particularly dimethyl disulfide, have proven biological activities against insects, and their use in insect pest management has been proposed as repellent [108].
Other BVOC-producing pathways are linked to biological matter processing, including the catabolism of fatty acids and lipid degradation, aromatic compound degradation, and protein and carbohydrate degradation. Catabolism of diet-introduced macromolecules by gut bacteria have a central role in the biosynthesis of semiochemicals such as those that are produced by the aromatic compound degradation pathways, i.e., benzaldehyde, benzoic acid, benzyl alcohol, guaiacol (2-methoxyphenol), phenol, phenylacetic acid, phenylethanol, phenylmethanol, and propionate. Many of these pathways have some intersections between them and, therefore, common intermediates. For example, propionate metabolism involves propionate and propanone, which is also relevant in the metabolism of ketone bodies, while aminobenzoate degradation involves guaiacol.
3.2. BVOCs and Secondary Metabolism
A number of specialized BVOC-producing pathways are linked to secondary metabolism. Bacterial secondary metabolism includes biochemical processes that, basically, are not essential for bacterial life, but play key roles in bacterial adaptation to unfavorable conditions, environmental response, and ecosystem regulation at the biodiversity level through species competition [109]. These functions are mostly encoded by genes outside the microbial “core genome” and are subject to intraspecies variability, although precursors of secondary metabolites come from primary metabolic pathways. Here, well-characterized or proposed pathways leading to three major classes of BVOCs that act as chemical signals in insects are illustrated. These classes include (I) pyrazines, (II) terpenes and terpenoids, and (III) alkanes and alkenes.
3.2.1. Pyrazine Metabolism in Bacteria
Pyrazines, an important class of BVOCs with chemical signaling properties in insects, are secondary metabolites that have been known since the early 1970s for their use in agriculture (insecticides and nematicides) or medicine (antidepressants, diuretics, and antibacterial) [110]. More recently, these compounds were described as flavor ingredients in the industries of chocolate and coffee [111], and also as anti-neoplastic [112] and anti-HIV drugs [113]. The pyrazine biosynthetic pathways exemplify how primary and secondary metabolisms are strictly interconnected. In general, in a secondary metabolism biosynthetic pathway, the first enzyme of a secondary biosynthetic pathway controls the metabolic flux, channeling precursors from the primary metabolites (amino acids, in pyrazine biosynthesis) toward specialized secondary metabolites. Thereafter, different sets of reactions occur for precursors assembling in the molecular skeleton and post-assembly modification.
Production of pyrazines for commercial purposes have allowed a better understanding of both the physiology of the pyrazine-producing microorganisms and the details of the biosynthetic pathways underlying their synthesis. Although there are many studies that have treated the biosynthesis and production of bacterial pyrazines as secondary metabolites, only few studies have reported their possible activity as semiochemicals in insects. We know, for instance, that 2,5-dimethylpyrazine is an important pheromone in ants Atta sexdens and it is produced by Citrobacter freundii [114] and Microbacterium foliorum [115]. In addition, 2,5-dimethyl-pyrazine and other pyrazines (2,3,5-trimethyl-pyrazine and tetramethyl-pyrazine) are produced by bacteria of the genus Bacillus by industrial fermentations [116]). Rajini and coworkers [117] reviewed some industrial applications of microbial pyrazines, the producer strains, and the proposed biosynthetic pathways.
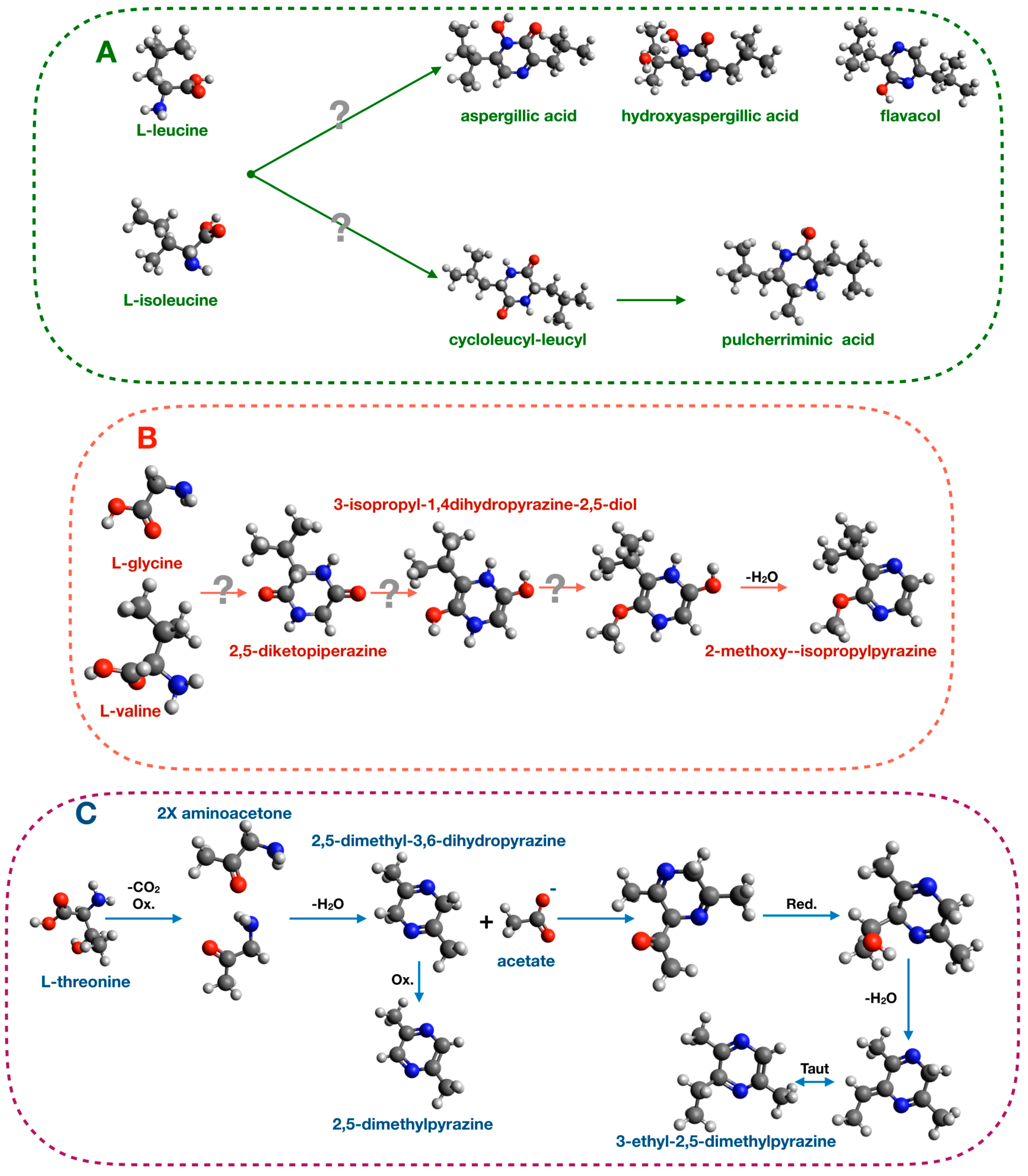
Many studies demonstrated that pyrazine biosynthesis is tightly linked to amino acid metabolism. For examples, 2,5-diisopropyl-pyrazine, 2-isopropyl-5-secbutyl-pyrazineand, and 17 other pyrazines were produced upon valine feeding of Paenibacillus polymyxa cultures [118]. These BVOCs are also produced by bacteria of the genus Exiguobacterium and act as oviposition signals in Anopheles gambiae [32,33]. In yeasts of the genus Aspergillus (Aspergillus flavus and Aspergillus oryzae), a similar experiment of amino acid feeding demonstrated that leucine and isoleucine supplementation of the culture medium enhanced the production of aspergillic acid [119]. This compound has antibacterial and chemoprotective activities [120,121]. Other pyrazines produced by the yeast of the genus Aspergillus (Aspergillus sclerotorium [122] and Aspergillus parasiticus [123]) are listed in Table S1. The proposed biosynthetic mechanism is based on condensation reaction between leucine and isoleucine and is depicted in Figure 4.
Another pyrazine, pulcherriminic acid, (2,5-diisobutyl-3,6-dihydroxypyrazine-1,4-dioxide) is synthesized by various bacteria, including Bacillus subtilis, Micrococcus violagabriellae, and Paenibacillus maceran, as well as by the yeast Candida pulcherrima [119,120,121,122,123,124,125,126,127]. In B. subtilis and C. pulcherrima, the proposed pathway for biosynthesis of this pyrazine produces a cyclic dipeptide cyclo l-leucyl-l-leucyl intermediate that is then converted in pulcherriminic acid [127]. Another proposed pathway in Pseudomonas perolens starts from valine and glycine to generate 2-methoxy-3-isopropylpyrazine [128] (Figure 4).
Several bacteria belonging to the Enterobacteriaceae family, particularly bacteria of the Serratia genus, are well-known for their ability to produce pyrazines. Specifically, Serratia oderifera, Serratia ficaria, and Serratia rubidea synthesize the alkylated pyrazines 3-isopropyl-2-methoxy-5-methylpyrazine, 3-isopropyl-2-methoxy-methylpyrazine, and 3-isobutyl-methoxy-pyrazine) [129]. A recent study demonstrated that L-trenonine and acetic acid are the precursors for 2,5-dimethylpyrazine and 3-ethyl-2,5-dimethylpyrazine synthesis in Serratia marcescens [37]. Radiolabeled precursors were used in this study and their incorporation into the final compounds was shown, leading to the proposed pathway illustrated in Figure 4.
Other microorganisms contributing to chemical signaling in insects as biotransforming agents include plant-derived pyrazines introduced by diet, which metabolized by some bacterial strains using two major metabolic pathways [117]. The first one converts hydroxy-pyrazines to glycine by oxidative degradation as reported in bacteria of the genus Pseudomonas [130], while the other one, which was reported in bacteria of the genus Stenotrophomonas, metabolizes hydroxy-pyrazines through an initial reduction, although the pathway is not well-characterized [131]. Another interesting example of pyrazine catabolism occurs in Mycobacterium tuberculosis. In vivo, this microorganism is sensitive to pyrazinamide, a first-line drug recommended by the World Health Organization for the treatment of tuberculosis in 1995. The drug, in vivo but not in vitro, is converted to pyrazinoic acid by pncA-encoded Mn2+/Fe2+ nicotinamidase [132]. Pyrazinoic acid inhibits FAS I (Fatty acids synthase I) and acidifies the microenvironment [133], causing the subsequent disruption of the membrane potential and disassembly of the membrane [134].
3.2.2. Terpene and Terpenoid Metabolism in Bacteria
Terpenes, terpenoids, and, more in general, isoprenoids, are a large class of compounds widespread among plants, yeasts, archaea, and bacteria with various functions, such as hormones [135], photosynthetic pigment [136], and major structural components in the archaeal membrane [137]. The isoprenoid class includes cholesterol and liposoluble vitamins A, D, E, and K [138]. In plants, many terpenes and terpenoids are multifunctional compounds. They protect plants against herbivores and pathogens, attract mutualists such as pollinators, and may act also as chemical signals for other plants or insects [139,140].
Different species of Dendroctonus are parasites of conifers, which are plants that produce various terpenes useful for protecting themselves from herbivorous insects [141]. The insect gut community affects tolerance to terpenes and terpenoids. Some bacterial strains metabolize these dangerous compounds, thereby reducing their toxicity and improving the insect survival [142]. On the other hand, in Dendroctonus valens multifunctional pheromone verbenone is synthesized by the gut bacterial community by biotransforming conifer monoterpenes ingested by diet [23,24], showing the promiscuous nature of these compounds.
Other terpenes act as hormones in plants, such as in the case of gibberellins, a class of diterpenes. In plants, gibberellins control development at different levels including seed germination, stem elongation, and flower induction. Plants, bacteria, and fungi synthesize these compounds, but the biosynthetic pathways are distinct in the different organisms. Plants use two diterpene synthases to form ent-kaurene, the giberellin precursor, from geranylgeranyl diphosphate (GGPP), while fungi use only a single bifunctional diterpene synthase. In both plants and fungi, ent-kaurene is then oxidized to ent-kaurenoic acid by cytochromes P450. ent-kaurenoic acid is further metabolized by different sets of enzymes in plants (sequentially: Two cytochromes P450, two 2-oxoglutarate dependent dioxygenases) and fungi (sequentially: Two cytochromes P450, desaturase, cytochrome P450) to obtain the final product, gibberellin A (GA3) [143]. Bacteria have evolved a distinct pathway, as shown in Bradyrhizobium japonicum, which contains an operon consisting of genes coding for a ferredoxin, a short-chain alcohol dehydrogenase, three cytochrome P450, a GGPP synthase, and two diterpene synthases (ent-copalyl diphosphate synthase and ent-kaurene synthase), which control giberellin biosynthesis.
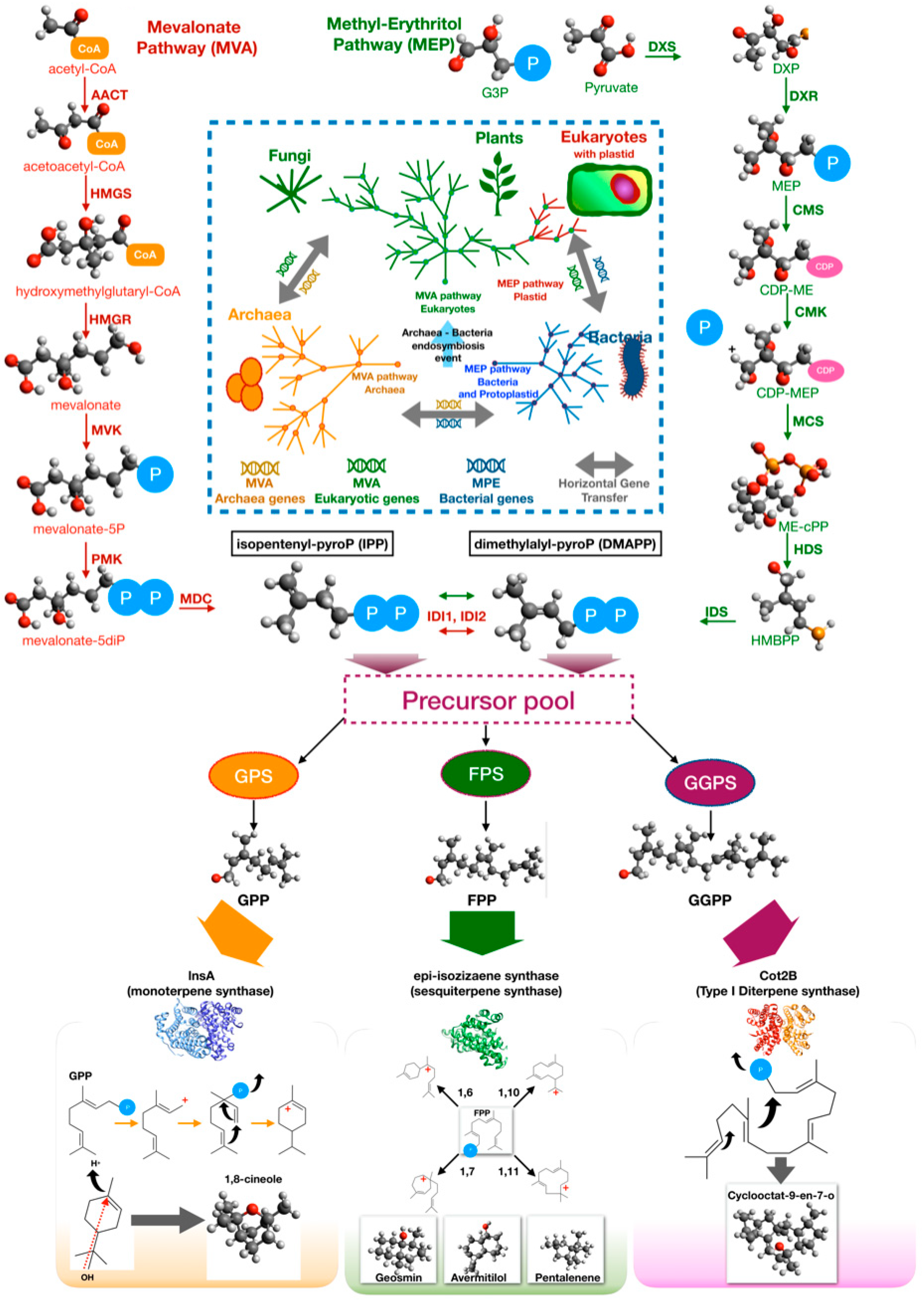
To synthesize terpenes and terpenoids, two interconvertible building blocks are required: Isopentenyl pyrophosphate (IPP) and dimethylallyl pyrophosphate (DMAPP). Biosynthesis of these precursors involves two pathways, i.e., the mevalonate pathway (MVA) and the non-mevalonate pathway, also known as 2-C-methyl-d-erythrol 4-phosphate (MEP) pathway [144,145]. The MVA pathway is considered as the fundamental pathway for isoprenoid biosynthesis in archaea and eukaryotes, although it seems to be present also in several bacteria. There are three hypotheses about the evolution of MVA pathway [145]. The first hypothesis contends that an ancestral pathway appeared in a common ancestor of eukaryotes and archaea, and it was recently acquired by few bacteria by horizontal genetic transfer (HGT) from a eukaryotic or archaeal donor. The second one proposes a similar model in which the HGT occurred in a common ancestor of a group of modern-day bacteria. The third hypothesis proposes that the MVA pathway was ancestral not only to archaea and eukaryotes, but also to bacteria. However, it was lost by most of bacteria. In this scenario, the last universal common ancestor life (LUCA) possessed the MVA pathway [145]. A number of Actinobacteria, Bacterioidetes, Firmicutes, Chloroflexi, Proteobacteria, and Spirochetes genomes contain homologs of MVA pathway genes, but not the entire classic pathway [145]. It has been proposed that the non-individuated MVA pathway enzymes likely evolve independently to complete the pathway, are supplementary enzymes that can be provided in trans configuration by other pathways, or are moonlighting proteins with two or more functions [146,147].
Details of the MVA pathway are illustrated in Figure 5. The first three steps, called the lower mevalonate pathway, are catalyzed, respectively, by acetoacetyl-CoA thiolase (AACT), HMG-CoA synthase (HMGS), and HMG-CoA reductase (HMGR) [148]. The result of lower pathway is the key precursor mevalonate and is then converted to mevalonate 5-phosphate by mevalonate kinase (MVK). This molecule is converted into IPP by two reactions which, in eukaryotes, involve phospho-mevalonate kinase (PMK) and mevalonate-diphosphate decarboxylase (MDC) while, in archaea, they involve a putative phospho-mevalonate decarboxylase (MPD) and a characterized isopentenyl phosphate kinase (IPK) [148]. Isomerization of IPP to DMAPP is carried out by distinct isopentenyl pyrophosphate isomerases in eukaryotes (IDI1) and archaea (IDI2) [148].
There is evidence that the MVA pathway exists and is essential in several bacteria, including staphylococci, streptococci, and enterococci, at variance with Bacillus subtilis and gram-negative bacteria that possess the MEP pathway. For example, in S. aureus, the mevalonate kinase was isolated and well-characterized [149]. Furthermore, knockout of the MVA pathway genes of Streptococcus pneumoniae stopped the growth in vitro under standard conditions. The growth was restored by adding mevalonate to the culture medium, demonstrating that this pathway is essential in this pathogenic bacterium and might represent a target for new antimicrobial compound [150].
The MEP is diffused in bacteria and plants. MVA and MEP coexist in eukaryotic organisms equipped with a plastid that evolved from a photosynthetic microorganism similar to modern-day cyanobacteria, according to the endosymbiotic theory [151]. These pieces of evidence suggest that MEP is characteristic of prokaryotes, and it was later acquired by eukaryotes with the photosynthetic endosymbiont, although different origins have been proposed [152].
Details of the MEP pathway are illustrated in Figure 5. The MEP pathway begins with the addition of pyruvate to glyceraldehyde-3-phosphate with the decarboxylation of pyruvate to yield 1-deoxy-d-xylulose-5-phosphate (DXP) and CO2. This reaction is catalyzed by DXP synthase (DXS) [153]. However, the same reaction occurs in E1 subunit of pyruvate kinase or decarboxylase as moonlighting activity [154,155]. The next steps are wel- characterized and are catalyzed by enzymes encoded by 6 isp genes: ispC, ispD (ygbP), ispE (ychB) and ispF (ygbB), ispG (gcpE), ispH (lytB) [153]. DXP is reduced to 2-C-methyl-d-erythritol 4-phosphate (MEP) by DXP reductoisomerase (IspC, DXR) [156,157]. MEP is converted 4-diphosphocytidyl-2-C-methyl-d-erythritol (CDP-ME) by CDP-ME synthase (IspD, CMS) using CTP [158]. CDP-ME kinase (IspE, CMK) phosphorylates CDP-ME yielding 4-diphosphocytidyl-2-C-methyl-d-erythritol 2-phosphate (CDP-MEP); 2-C-methyl-d-erythritol 2,4-cyclodiphosphate synthase (IspF, MCS), then detaches the CMP and converts the intermediate into phosphocyclic compound 2-C-methyl-d-erythritol 2,4-cyclodiphosphate (ME-cPP) [159,160]. Finally, hydroxy-methyl-butenyl 4-diphosphate [HMBPP] synthase (IspG, HDS) catalyzes the synthesis of HMBPP from ME-cPP, while 4-hydroxy-3-methylbut-2-enyl-diphosphate reductase (IspH, HDR) synthesizes a mixture of IPP and DMAPP [161]. In the MEP pathway, as well as in the MVA pathway, an isomerase (IDI) catalyzes the interconversion of IPP and DMAPP [162].
Although all terpenes are synthesized from two common precursors, the basic C5 isoprene units IPP and DMAPP, the structural and functional diversity of the final products are surprisingly high. Such a diversity is largely dependent on the biosynthetic enzymes that process the precursors to form the terpene rings [143]. The head-to-tail condensation of IPP to DMAPP results in the formation of geranyl pyrophosphate (GPP), and the further addition of another IPP unit leads to production of farnesyl pyrophosphate (FPP). Geranylgeranyl pyrophosphate is formed by the condensation of one FPP with one IPP unit. GPP, FPP, and GGPP are, respectively, the acyclic precursors for the biosynthesis of monoterpene, sesquiterpene, and diterpene, which are the most common terpenes in bacteria (Figure 5). Cyclization reactions are catalyzed, respectively, by monoterpene cyclases, sesquiterpene cyclases, and diterpene cyclases.
The first bacterial monoterpene synthase (encoded by cnsA) was isolated from Streptomyces clavuligerus ATCC 27064 by Ohnishi and coworkers [163]. This enzyme catalyzes the biosynthesis of 1,8-cineole from GPP involving an α-terpineol intermediate until the complete conversion. The reference monoterpene synthase, 2-methylisoborneol synthase, instead uses (E)-2-methyl-GPP as a substrate. The product, 2-methylisoborneol, is volatile homoterpene alcohol with an earthy, musty odor. The enzyme mechanism is well-understood and involves ionization of the substrate followed by isomerization and final cyclization [164]. A second monoterpene synthase was identified in another Streptomyces clavuligerus, the linalool/nerolidol synthase (lnsA). This enzyme catalyzes the synthesis of the acyclic terpenoids (3R)-linalool and (3R)-(E)-nerolidol using, GPP and FPP as substrates, respectively [165]. Finally, as a moonlighting function, several sesquiterpene synthases may also produce monoterpenes using GPP, an unusual substrate for this type of terpene synthase. This hypothesis was demonstrated by heterologous expression in E. coli of the epicubebol synthase from Streptosporangium roseum [166].
Sesquiterpenes synthases are the major class of terpene synthases that are found in bacteria. These enzymes use FPP as substrate, but different enzymes can cyclize the precursors with different mechanisms, resulting in various ring closures: 1,6 (bisabolyl catione), 1,7 (cycloheptenyl cation), 1,10 ((E,E)-germacrenyl cation), and 1,11 ((E,E)-humulyl cation). Usually, when an ionic intermediate is involved in the biosynthetic pathways, an enantiomer mixture may result from the reaction, thereby expanding the repertoire of synthesized molecules [167]. A typical sesquiterpene synthase is pentalenene synthase from Streptomyces exfoliates, an enzyme related to the synthase of pentalenolactone, which was isolated from Streptomyces roseogriseus in 1957 as an antibiotic [168]. Geosmin synthase is another common sesquiterpene synthase in streptomycetes. Geosmin (l,2,7,7-tetramethyl-2-norborneol) is a terpene compound that is responsible for the earthy scent released when rain falls on dry soil. The gene encoding this biosynthetic activity was first identified in the model organism Streptomyces coelicolor A3(2) [169]. It is interesting to note that 2-methylisoborneol and geosmin, two rather common BVOCs produced by many streptomycetes, have powerful biological effects against insects. A system and method for attracting ants by geosmin and/or repelling ants by 2-methylisoborneol was patented [170].
Diterpene synthases are a complex and relatively misunderstood class of enzymes that catalyze the conversion of GGPP into diterpene molecules. Two types of diterpene synthases (type I and II) can be distinguished. Type I diterpene synthases transform directly the substrate into the final terpenes, while type II diterpene synthases catalyzes the synthesis of diphosphate intermediates that can be processed by type I cyclases. The reference enzyme for type I diterpene synthases is cyclooctat-9-en-7-ol synthase (CotB2), which was isolated from Streptomyces melanosporofaciens MI614-43F2. This enzyme is involved in the biosynthesis of cyclooctatin, a lysophospholipase inhibitor [171,172]. The first bacterial type II diterpene synthase reported in literature was the terpentedienyl diphosphate synthase from Streptomyces griseolosporus MF730-N6, which catalyzes the synthesis of terpentedienyl diphosphate that is converted into terpentetriene, the precursor of the antibiotic terpentecin [173,174].
3.2.3. Alkane and Alkene Metabolism in Bacteria
CHCs play a central role in social insect communication and influence various aspects of their behavior and health. Many studies have focused on CHCs as both pheromones and signaling molecules. However, few studies have addressed their biosynthetic pathways, which therefore remain poorly characterized. Initial studies identified the biosynthetic source of CHCs in the insects’ cells only, but further studies demonstrated the relevance of the microbiota metabolism as described above. Many studies report the ability of certain groups of bacteria to synthesize linear alkanes and alkenes, but even in this case, the biosynthetic pathways remain poorly characterized, with some exceptions. Most of the information comes from studies on cyanobacteria [175].
Different biosynthetic pathways leading to alkanes and/or alkenes from fatty acid precursors were proposed: (I) the Acyl-ACP reductase-aldehyde-deformylating oxygenase pathway (AAR-ADO), (II) the olefin biosynthetic pathway (Ole, encoded by the oleABCD operon), (III) the terminal olefin pathway (OleTJE), (IV) the olefin synthase pathway (Ols) (Figure 6). Of all these pathways, only the first two (AAR-ADO and Ole) are well-characterized [175].
The AAR-ADO was first identified in Synechococcus [176] using a differential and subtractive genomic approach based on the differences between alkane/alkene producing and nonproducing strains. Evidence was provided that alkanes and alkenes biosynthesis start from an acyl-acylated acyl-carrier protein (ACP). The acyl-ACP substrate is then reduced by acyl-ACP reductase (AAR) and the resulting aldehyde is converted to terminal alkane or alkene by aldehyde-deformylating oxygenase (ADO) [177]. ADO is a unique enzyme of cyanobacteria, which converts acyl aldehyde precursors into alkane/alkene by the combined action of ferredoxin and NADPH:ferredoxin/flavodoxin oxidoreductase and O2 as an electron acceptor [177].
The Ole pathway was characterized in Micrococcus luteus [167]. The M. luteus olefin biosynthetic operon oleABCD is composed of three genes that codes for enzymes: Thiolase (OleA), bifunctional α/β hydrolase and AMP-dependent ligase/synthetase (OleB/C), and short-chain dehydrogenase (OleD) [178]. The pathway starts with a nondecarboxylative Claisen condensation catalyzed by OleA, which generates a β-keto acid that is then reduced by OleD in a NADPH-dependent reaction to produce a β-hydroxy acid. OleC catalyzes the conversion of the β-hydroxy acid to an alkene using ATP. OleB is thought to perform scaffolding or regulatory function in the Ole complex. This pathway generates nonterminal long-chain alkenes by fatty acid head-to-head condensation.
Terminal olefins are, instead, produced by Jeotgalicoccus using OleTJE, a fatty acid decarboxylase that was annotated as P450 peroxygenase of cyp152 family. OleTJE uses H2O2 as an electron source to decarboxylate fatty acids releasing H2O and CO2 [179].
The Ols pathway was described in Synechococcus. In this photosynthetic cyanobacterium, the olefin synthase (CurM), a multifunctional protein very similar to polyketide synthases (PKSs), was characterized. As well as bacterial PKS involved in the synthesis of antibiotics, CurM is composed of multiple domains: A loading domain (LD), two ACP domains (ACP1 and ACP2), a central ketosynthase (KS) domain, an acyltransferase (AT) domain, a ketoreductase (KR), a sulfotransferase (ST) domain, and a carboxy-terminal thioesterase (TE) domain [180].
In addition to the example related to pyrazines, biotechnological exploitation of the ability of certain bacteria to produce alkanes/alkenes is leading to a better understanding of the underlying biosynthetic pathways. An artificial pathway (CAR/FAR-ADO) was implemented in E. coli to produce precursor (aldehydes) using (I) carboxylic acid reductase (CAR) from Mycobacterium marinum, (II) phosphopantetheinyl transferase Sfp from Bacillus subtilis, and (III) fatty acid reductase (FAR) complex (LuxC, LuxE, and LuxD) from Photorhabdus luminescens. This precursor was converted by ADO to produce hydrocarbons [181,182].
Although alkanes and alkenes can be produced by bacteria, insects can contribute to their synthesis. For example, in Drosophila, one enzyme (P450) of the CYP4G family oxidatively produces hydrocarbons, starting from aldehydes precursors [183].
4. Conclusions and Perspectives
Despite the involvement of microorganisms, the chemical communication between insects and plants is now recognized due to numerous studies. However, many questions still remain unanswered. Many of these concerns arise from the fact that the majority of studies in this research area were mostly observational. They allow us to associate certain phenotypes (or behaviors) in insects with a specific structure of the insect microbial community, but they do not allow us to detect the cause–effect relationships. On the other hand, experimental in vitro studies are often limited in the validity of the system or in the analysis model. Very few experimental studies are carried out in the natural environment.
In other words, in many cases, there is very clear observational evidence regarding the role played by several microorganisms in such transkingdom interactions. In some cases, there is also clear evidence that these microorganisms can produce in vitro molecules that may act as chemical signals in insects and/or plants. On the contrary, in most cases, formal proofs are often missing (I) that there is production of these bioactive molecules in the natural environment where these microorganisms live, and (II) that their amount is sufficient to elicit a biological response in the host. The measurement of microbial activity in the natural environment is very difficult, especially when it is necessary to quantify compounds that are produced in limited amounts during the microorganism–host interaction and may be produced also by the host. Most of the results of the studies reported here must be considered in light of these limitations, despite the attempt to address this point, to put together ecological and available biochemical information.
Nevertheless, there is additional evidence supporting that the hypothesis that BVOCs play a role in transkingdom interactions and are not simply byproducts of bacterial primary or secondary metabolism. This evidence comes from gene regulation studies. On the one hand, it is expected that the production of BVOCs is influenced by chemical–physical parameters, such as nutrient availability, temperature, pH, relative humidity, and the presence or absence of oxygen. On the other hand, recent studies have shown that the production of BVOCs can be stimulated or inhibited during the interaction of bacteria with their hosts consistently with a biological activity of BVOCs in the host [184,185,186,187]. Specifically, in several bacteria, BVOCs production is controlled by GacS/GacA two-component system which, in enteric bacteria and fluorescent pseudomonads, controls primary and energy metabolism and the expression of virulence (secretion systems) or biocontrol factors, including exoenzymes and secondary metabolites (siderophores, antibiotics, BVOCs) [187,188,189].
Other questions still awaiting an answer concern the BVOCs biosynthetic pathways that are, for many BVOCs, still not well-characterized. Here, we attempted to call attention to some biosynthetic pathways of primary importance in the transkingdom interaction between bacteria, insects, and plants, which lead to the production of pyrazines, terpenes and terpenoids, alkanes, and alkenes. We highlighted how some information about the biosynthetic pathways comes from researches in allied fields, such as those investigating the biotechnological exploitation of all these compounds. The elucidation of the biosynthetic pathways underlying BVOCs production will not only will help us better understand the biological role of these compounds, but it will also facilitate their use in various fields, from agriculture to food-processing and food-control, from medicine to veterinary, and from energy production (as biofuels) to environmental management.
Supplementary Materials
The following are available online at https://www.mdpi.com/2075-4450/10/12/441/s1, Table S1. Insect semiochemicals produced by microorganisms. All molecules that were cited in this review were listed here. Columns: (A) IUPAC name of semiochemical compound; (B) alternative name of semiochemical compound; (C) chemical/biosynthetic class of semiochemical compound; (D–F) Order (D), Family (E), Genus and species (F) of insect arthropods that are targeted by semiochemical compound; (G–I) Order (G), Family (H), Genus and species of microorganisms producing semiochemical (I); (J,K) source of isolation of microorganism (J), role of microorganisms in semiochemical metabolism (K); (L,M) general function (L) and specific function (M) of semiochemical compound; (N,O) reference number (N) and abbreviated reference (O); (P–R) code of compound in KEGG database (hyperlink with KEGG) (P), related KEGG pathway map (Q), related KEGG pathway name (R); (S–U) chemical class of semiochemical compound as indicated in Pherobase, hyperlinks for information found in Pherobase: compound with behavioral function (T) or compound without assigned function (U).
Author Contributions
Conceptualization, P.A.; Data acquisition and processing, P.A., M.C.; writing-review and editing, P.A., M.C., A.T., S.M.T.
Funding
This work was partially supported by project PON BIOMIS ARS01_01220.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nihorimbere, V.; Ongena, M.; Smargiassi, M.; Thonart, P. Beneficial effect of the rhizosphere microbial community for plant growth and health. Biotechnol. Agron Soc. Environ. 2011, 15, 327–337. [Google Scholar]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial Services of Arbuscular Mycorrhizal Fungi—From Ecology to Application. Front. Plant Sci. 2018, 9, 1270. [Google Scholar] [CrossRef] [PubMed]
- Moreau, D.; Bardgett, R.D.; Finlay, R.D.; Jones, D.L.; Philippot, L. A plant perspective on nitrogen cycling in the rhizosphere. Funct. Ecol. 2019, 3, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, D.K.; Kasotia, A.; Jain, S.; Vaishnav, A.; Kumari, S.; Sharma, K.P.; Varma, A. Bacterial-Mediated Tolerance and Resistance to Plants Under Abiotic and Biotic Stresses. J. Plant Growth Regul. 2016, 35, 276–300. [Google Scholar] [CrossRef]
- Heil, M.; Bostock, R.M. Induced Systemic Resistance (ISR) Against Pathogens in the Context of Induced Plant Defences. Ann. Bot. 2002, 89, 503–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.K.; McInroy, J.A.; Kloepper, J.W. The Interactions of Rhizodeposits with Plant Growth-Promoting Rhizobacteria in the Rhizosphere: A Review. Agriculture 2019, 9, 142. [Google Scholar] [CrossRef] [Green Version]
- Vavre, F.; Kremer, N. Microbial impacts on insect evolutionary diversification: From patterns to mechanisms. Curr. Opin Insect Sci. 2014, 4, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Pineda, A.; Zheng, S.; van Loon, J.J.A.; Pieterse, C.M.J.; Dicke, M. Helping plants to deal with insects: The role of beneficial soil-borne microbes. Trends Plant Sci. 2010, 15, 507–514. [Google Scholar] [CrossRef]
- Ueda, K.; Tawaraya, K.; Murayama, H.; Sato, S.; Nishizawa, T.; Toyomasu, T.; Murayama, T.; Shiozawa, S.; Yasuda, H. Effects of arbuscular mycorrhizal fungi on the abundanceof foliar-feeding insects and their natural enemy. Appl. Entomol. Zool. 2013, 48, 79–85. [Google Scholar] [CrossRef]
- Hempel, S.; Stein, C.; Unsicker, S.B.; Renker, C.; Auge, H.; Weisser, W.W.; Buscot, F. Specific bottom–up effects of arbuscular mycorrhizal fungi across a plant–herbivore–parasitoid system. Oecologia 2009, 160, 267. [Google Scholar] [CrossRef] [Green Version]
- Zebelo, S.A.; Bertea, C.M.; Bossi, S.; Occhipinti, A.; Gnavi, G.; Maffei, M.E. Chrysolina herbacea Modulates Terpenoid Biosynthesis of Mentha aquatica L. PLoS ONE 2011, 6, e17195. [Google Scholar] [CrossRef] [Green Version]
- Engl, T.; Kaltenpot, M. Influence of microbial symbionts on insect pheromones. Nat. Prod. Rep. 2018, 35, 386. [Google Scholar] [CrossRef] [PubMed]
- Leroy, P.D.; Sabri, A.; Verheggen, F.J.; Francis, F.; Thonart, P.; Haubruge, E. The semiochemically mediated interactions between bacteria and insects. Chemoecology 2011, 21, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Van der Meer, R. Ant Interactions with Soil Organisms and Associated Semiochemicals. J. Chem. Ecol. 2012, 38, 728–745. [Google Scholar] [CrossRef] [PubMed]
- Wielkopolan, B.; Obrępalska-Stęplowska, A. Three-way interaction among plants, bacteria, and coleopteran insects. Planta 2016, 244, 313–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kather, R.; Martin, S.J.J. Evolution of Cuticular Hydrocarbons in the Hymenoptera: A Meta-Analysis. Chem. Ecol. 2015, 41, 871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schettino, M.; Grasso, D.A.; Weldegergis, B.T.; Castracani, C.; Mori, A.; Dicke, M.; Van Lenteren, J.C.; Van Loon, J.J.A. Response of a Predatory ant to Volatiles Emitted by Aphid-and Caterpillar-Infested Cucumber and Potato Plants. J. Chem. Ecol. 2017, 43, 1007. [Google Scholar] [CrossRef]
- Beck, J.J.; Alborn, H.T.; Block, A.K.; Christensen, S.A.; Hunter, C.T.; Rering, C.C.; Seidl-Adams, I.; Stuhl, C.J.; Torto, B.; Tumlinson, J.H. Interactions Among Plants, Insects, and Microbes: Elucidation of Inter-Organismal Chemical Communications in Agricultural Ecology. J. Agric. Food Chem. 2018, 66, 6663–6674. [Google Scholar] [CrossRef]
- Jordan, H.R.; Tomberlin, J.K. Abiotic and Biotic Factors Regulating Inter-Kingdom Engagement between Insects and Microbe Activity on Vertebrate Remains. Insects 2017, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial Volatile Emissions as Insect Semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef]
- Dillon, R.; Charnley, K. Mutualism between the desert locust Schistocerca gregaria and its gut microbiota. Res. Microbiol. 2002, 153, 503–509. [Google Scholar] [CrossRef]
- Dillon, R.; Vennard, C.T.; Charnley, A.K. Exploitation of gut bacteria in the locust. Nature 2000, 403, 851. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Lou, Q.; Cheng, C.; Lu, M.; Sun, J. Gut-Associated Bacteria of Dendroctonus valens and their Involvement in Verbenone Production. Microb. Ecol. 2015, 70, 1012–1023. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.S.; Boone, C.K.; Bohlmann, J.; Raffa, K.F. Responses of Bark Beetle-Associated Bacteria to Host Monoterpenes and Their Relationship to Insect Life Histories. J. Chem. Ecol. 2011, 37, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Boone, C.K.; Keefover-Ring, K.; Mapes, A.C.; Adams, A.S.; Bohlmann, J.; Raffa, K.F. Bacteria Associated with a Tree-Killing Insect Reduce Concentrations of Plant Defense Compounds. J. Chem. Ecol. 2013, 39, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Wada-Katsumata, A.; Zurekc, L.; Nalyanya, G.; Roelofs, W.L.; Zhangf, A.; Schala, C. Gut bacteria mediate aggregation in the German cockroach. Proc. Natl. Acad. Sci. USA 2015, 112, 15678–15683. [Google Scholar] [CrossRef] [Green Version]
- Uriel, Y.; Gries, R.; Tu, L.; Carroll, C.; Zhai, H.; Moore, M.; Gries, G. The fly factor phenomenon is mediated by interkingdom signaling between bacterial symbionts and their blow fly hosts. Insect Sci. 2018, 1–10. [Google Scholar] [CrossRef]
- Verheggen, F.J.; Diez, L.; Sablon, L.; Fischer, C.; Bartram, S.; Haubruge, E.; Detrain, C. Aphid alarm pheromone as a cue for ants to locate aphid partners. PLoS ONE 2012, 7, e41841. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.Y.; Lognay, G.C.; Detrain, C.; Heil, M.; Grigorescu, A.; Sabri, A.; Thonart, P.; Haubruge, E.; Verheggen, F.J. Bacteria may enhance species association in an ant–aphid mutualistic relationship. Chemoecology 2015, 25, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.Y.; Detrain, C.; Thonart, P.; Haubruge, E.; Francis, F.; Verheggen, J.F.; Lognay, G.C. Bacteria may contribute to distant species recognition in ant–aphid mutualistic relationships. Insect Sci. 2017, 24, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Francis, F.; Vandermoten, S.; Verheggen, F.; Lognay, G.C.; Haubruge, E. Is the (E)-β-farnesene only volatile terpenoid in aphids? J. Appl. Entomol. 2005, 129, 6–11. [Google Scholar] [CrossRef]
- Sumba, L.A.; Guda, T.O.; Deng, A.L.; Hassanali, A.; Beier, J.C.; Knols, B.G.J. Mediation of oviposition site selection in the African malaria mosquito Anopheles gambiae (Diptera: Culicidae) by semiochemicals of microbial origin. Int. J. Trop. Insect Sci. 2004, 24, 260–265. [Google Scholar] [CrossRef]
- Lindh, J.M.; Kännaste, A.; Knols, B.G.J.; Faye, I.; Borg-Karlson, A.K. Oviposition Responses of Anopheles gambiae s.s. (Diptera: Culicidae) and Identification of Volatiles from Bacteria-Containing Solutions. J. Med. Entomol. 2008, 45, 45–1039. [Google Scholar] [CrossRef] [Green Version]
- Verhulst, N.O.; Qiu, Y.T.; Beijleveld, H.; Maliepaard, C.; Knights, D.; Schulz, S.; Berg-Lyons, D.; Lauber, C.L.; Verduijn, W.; Haasnoot, G.W.; et al. Composition of Human Skin Microbiota Affects Attractiveness to Malaria Mosquitoes. PLoS ONE 2011, 6, e28991. [Google Scholar] [CrossRef]
- Verhulst, N.O.; Beijleveld, H.; Knols, B.G.J.; Takken, W.; Schraa, G.; Bouwmeester, H.J.; Smallegange, R.C. Cultured skin microbiota attracts malaria mosquitoes. Malar. J. 2009, 8, 302d. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Crippen, T.L.; Coates, C.J.; Wood, T.K.; Tomberlin, J.K. Effect of Quorum Sensingby Staphylococcus epidermidis on the Attraction Response of Female Adult Yellow Fever Mosquitoes, Aedes aegypti aegypti (Linnaeus) (Diptera:Culicidae), to a Blood-Feeding Source. PLoS ONE 2015, 10, e0143950. [Google Scholar] [CrossRef] [Green Version]
- Silva-Junior, E.A.; Ruzzini, A.C.; Paludo, C.R.; Nascimento, F.S.; Currie, C.R.; Clardy, J.; Pupo, M.T. Pyrazines from bacteria and ants: Convergent chemistry within an ecological niche. Sci. Rep. 2018, 8, 2595. [Google Scholar] [CrossRef] [Green Version]
- Babcock, T.; Borden, J.H.; Gries, R.; Carroll, C.; Lafontaine, J.P.; Moore, M.; Gries, G. Inter-kingdom signaling—Symbiotic yeasts produce semiochemicals that attract their yellowjacket hosts. Entomol. Exp. Appl. 2019, 167, 220–230. [Google Scholar] [CrossRef]
- Matsuura, K. Nestmate recognition mediated by intestinal bacteria in a termite, Reticulitermes speratus. Oikos 2001, 92, 20–26. [Google Scholar] [CrossRef]
- Nalepa, C.A. Origin of termite eusociality: Trophallaxis integrates the social, nutritional, and microbial environments. Ecol. Entomol. 2015, 40, 323–335. [Google Scholar] [CrossRef]
- Guo, L.; Quilici, D.R.; Chase, J.; Blomquis, G.J. Gut tract microorganisms supply the precursors for methyl-branched hydrocarbon biosynthesis in the termite, Zootermopsis nevadensis. Insect Biochem. 1991, 21, 327–333. [Google Scholar] [CrossRef]
- Teseo, S.; van Zweden, J.S.; Pontieri, L.; Kooij, P.W.; Sørensen, S.J.; Wenseleers, T.; Poulsen, M.; Boomsma, J.J.; Sapountzis, P. The scent of symbiosis: Gut bacteria may affect social interactions in leaf-cutting ants. Anim. Behav. 2019, 150, 239–254. [Google Scholar] [CrossRef]
- Powell, J.E.; Martinson, V.G.; Urban-Mead, K.; Moran, N.A. Routes of Acquisition of the Gut Microbiota of the Honey Bee Apis mellifera. Appl. Environ. Microbiol. 2014, 80, 7378–7387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, J.E.; Eiri, D.; Moran, N.A.; Rangel, J. Modulation of the honey bee queen microbiota: Effects of early social contact. PLoS ONE 2018, 13, e0200527. [Google Scholar] [CrossRef]
- Yun, J.H.; Jung, M.J.; Kim, P.S.; Bae, J.W. Social status shapes the bacterial and fungal gut communities of the honey bee. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kapheim, K.M.; Rao, V.D.; Yeoman, C.J.; Wilson, B.A.; White, B.A.; Goldenfeld, N.; Robinson, G.E. Caste-specific dif- ferences in hindgut microbial communities of honey bees (Apis mellifera). PLoS ONE 2015, 10, e0123911. [Google Scholar] [CrossRef]
- Tarpy, D.R.; Mattila, H.R.; Newton, I.L.G. Development of the honey bee gut microbiome throughout the queen-rearing process. Appl. Environ. Microbiol. 2015, 81, 3182–3191. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.C.; Fruciano, C.; Hildebrand, F.; Al Toufalilia, H.; Balfour, N.J.; Bork, P.; Engel, P.; Ratnieks, F.L.W.; Hughes, W.O.H. Gut microbiota composition is associated with environmental landscape in honey bees. Ecol. Evol. 2018, 8, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, D.; Narita, S.; Watanabe, M. Insect Sex Determination Manipulated by Their Endosymbionts: Incidences, Mechanisms and Implications. Insects 2012, 3, 161–199. [Google Scholar] [CrossRef] [Green Version]
- Burand, J.P.; Tan, W.J. Mate preference and mating behavior of male Helicoverpa zea (Lepidoptera: Noctuidae) infected with the sexually transmitted insect virus Hz-2V. Ann. Entomol. 2006, 99, 969–973. [Google Scholar] [CrossRef]
- Wigul, G.; Sikorowski, P.P. The Effect of Staphylococcal Enterotoxin B on Pheromone Production in Fat Bodies Isolated from Male Boll Weevils. J. Invertebr. Pathol. 1986, 47, 116–119. [Google Scholar] [CrossRef]
- Hoyt, C.P.; Osborne, G.O.; Mulcock, A.P. Production of an insect sex attractant by symbiotic bacteria. Nature 1971, 230, 472–473. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.G.; Jackson, T.A.; Unelius, C.R.; Wee, S.L.; Young, S.D.; Townsend, R.J.; Suckling, D.M. Morganella morganii bacteria produces phenol as the sex pheromone of the New Zealand grass grub from tyrosine in the colleterial gland. Sci. Nat. 2016, 103, 59. [Google Scholar] [CrossRef] [PubMed]
- Cremer, S.; Lautenschläger, B.; Heinze, J. A transitional stage between the ergatoid and winged male morph in the ant Cardiocondyla obscurior. Insectes Soc. 2002, 49, 221–228. [Google Scholar] [CrossRef]
- Heinze, J. Life-history evolution in ants: The case of Cardiocondyla. Proc. R Soc. 2017, B284, 20161406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, A.; Schrader, L.; Gil, R.; Manzano-Marin, A.; Florez, L.; Wheeler, D.; Werren, J.H.; Latorre, A.; Heinze, J.; Kaltenpoth, M.; et al. A novel intracellular mutualistic bacterium in the invasive ant Cardiocondyla obscurior. ISME J. 2016, 10, 376–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzolante, G.; Cordero, C.; Tredici, S.M.; Vergara, D.; Pontieri, P.; Del Giudice, L.; Capuzzo, A.; Rubiolo, P.; Kanchiswamy, C.N.; Zebelo, S.A.; et al. Cultivable gut bacteria provide a pathway for adaptation of Chrysolina herbacea to Mentha aquatica volatiles. BMC Plant Biol. 2017, 17, 30. [Google Scholar] [CrossRef] [Green Version]
- Maffei, M.E.; Chialva, F.; Sacco, T. Glandular trichomes and essential oil in developing peppermint leaves. I. Variation in peltate trichome number and terpene distribution within leaves. New Phytol. 1989, 111, 707–716. [Google Scholar]
- Wood, W.F.; Palmer, T.M.; Stanton, M.L. A comparison of volatiles in mandibular glands from three Crematogaster ant symbionts of the whistling thorn acacia. Biochem. Syst. Ecol. 2002, 30, 217–222. [Google Scholar] [CrossRef]
- El-Sayed, A.M. The Pherobase: Database of Pheromones and Semiochemicals. Available online: http://www.pherobase.com (accessed on 10 November 2019).
- Whitehead, S.R.; Reid, E.; Sapp, J.; Poveda, K.; Royer, AM.; Posto, A.L.; Kessler, A. A Specialist Herbivore Uses Chemical Camouflage to Overcome the Defenses of an Ant-Plant Mutualism. PLoS ONE 2014, 9, e102604. [Google Scholar] [CrossRef]
- Oliver, T.H.; Mashanova, A.; Leather, S.R.; Cook, J.M.; Jansen, V.A.A. Ant semiochemicals limit apterous aphid dispersal. Proc. R. Soc. B 2007, 274, 3127–3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, P.D.; Sabri, A.; Heuskin, S.; Thonart, P.; Lognay, G.; Verheggen, F.J.; Francis, F.; Brostaux, Y.; Felton, G.W.; Haubruge, E. Microorganisms from aphid honeydew attract and enhance the efficacy of natural enemies. Nat. Commun. 2011, 2, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, N.E.; Braby, M.F.; Heath, A.; Lohman, D.J.; Mathew, J.; Rand, D.B.; Travassos, M.A. The ecology and evolution of ant association in the Lycaenidae (Lepidoptera). Annu. Rev. Entomol. 2002, 47, 733–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Als, T.D.; Nash, D.R.; Boomsma, J.J. Adoption of parasitic Maculinea alcon caterpillars (Lepidoptera:Lycaenidae) by three Myrmica ant species. Anim. Behav. 2001, 62, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Barbero, F. Cuticular Lipids as a Cross-Talk among Ants, Plants and Butterflies. Int. J. Mol. Sci. 2016, 17, 1966. [Google Scholar] [CrossRef] [Green Version]
- Di salvo, M.; Calcagnile, M.; Talà, A.; Tredici, S.M.; Maffei, M.E.; Schönrogge, K.; Barbero, F.; Alifano, P. The Microbiome of the Maculinea-Myrmica Host-parasite Interaction. Sci. Rep. 2019, 9, 8048. [Google Scholar] [CrossRef] [Green Version]
- Szenteczki, M.A.; Pitteloud, C.; Casacci, L.P.; Kešnerová, L.; Whitaker, M.R.L.; Enge, P.; Vila, R.; Alvarez, N. Bacterial communities within Phengaris (Maculinea) alcon caterpillars are shifted following transition from solitary living to social parasitism of Myrmica ant colonies. Ecol. Evol. 2019, 9, 4452–4464. [Google Scholar] [CrossRef] [Green Version]
- Kronauer, D.J.C.; Pierce, N.E. Myrmecophiles. Curr. Biol. 2011, 21, 6. [Google Scholar] [CrossRef] [Green Version]
- Tsuchida, T.; Koga, R.; Horikawa, M.; Tsunoda, T.; Maoka, T. Symbiotic bacterium modifies aphid body color. Science 2010, 330, 1102–1104. [Google Scholar] [CrossRef]
- Menzel, R.; Backhaus, W. Colour vision in insects. In Vision and Visual Dysfunction; Gouras, P., Ed.; Mac-Millan: London, UK, 1991; pp. 262–293. [Google Scholar]
- Greenfield, J.C. Vetiver Grass: The Hedge Against Erosion; World Bank: Washington, DC, USA, 1990; ISBN 10: 082131405X. [Google Scholar]
- Babalola, O.; Oshunsanya, S.O.; Are, K. Effects of vetiver grass (Vetiveria nigritana) strips, vetiver grassmulch and an organomineral fertilizer on soil, water andnutrient losses and maize (Zea mays, L.) yields. Soil Tillage Res. 2007, 96, 6–18. [Google Scholar] [CrossRef]
- Danh, L.T.; Truong, P.; Mammucari, R.; Foster, N. Economic Incentive for Applying Vetiver Grass to Remediate Lead, Copper and Zinc Contaminated Soils. Int. J. Phytoremed. 2010, 13, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Brandt, R.; Merkl, N.; Schultze-Kraft, R.; Infante, C.; Broll, G. Potential of vetiver (Vetiveria zizanioides (L.) Nash) for phytoremediation of petroleum hydrocarbon-contaminated soils in Venezuela. Int. J. Phytoremed. 2006, 8, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E. Vetiveria: The genus Vetiveria; Taylor & Francis Inc.: New York, NY, USA, 2002; ISBN 0-203-27411-3. [Google Scholar]
- Yoon, P.K. Extracts from A Look-See at Vetiver Grass in Malaysia-First Progress Report. Vetiver Newsl. 1991, 6, 86–96. [Google Scholar]
- Zisong, W. Excerpts from the experiments and popularization of vetiver grass, Nanpang Prefecture. Vetiver Newsl. 1991, 6, 105–108. [Google Scholar]
- Van den Berg, J. Vetiver grass (Vetiveria zizanioides (L.) Nash) as trap plant for Chilo partellus (Swinhoe) (Lepidoptera: Pyralidae) and Busseola fusca (Fuller) (Lepidoptera: Noctuidae). Ann. Soc. Entomol. Fr. 2006, 42, 449–454. [Google Scholar] [CrossRef]
- Van den Berg, J. Can Vetiver Grass be Used to Manage Insect Pests on Crops? In Proceedings of the Third Internatioal Vetiver Conference, Guangzhou, China, 6–9 October 2003. [Google Scholar]
- Chomchalow, N. Vetiver: A Living Trap; PRVN Tech. Bull.No. 2015/2; ORDPB: Bangkok, Thailand, 2015. [Google Scholar]
- Jain, S.C.; Nowicki, S.; Eisner, T.; Meinnald, J. Insect repellents from vetiver oil: Zizanal and epizizanal. Tetrahedron Lett. 1982, 23, 4639–4642. [Google Scholar] [CrossRef]
- Zhu, B.C.; Henderson, G.; Chen, F.; Maistrello, L.; Laine, R.A. Nootkatone is a repellent for Formosan subterranean termite (Coptotermes formosanus). J. Chem. Ecol. 2001, 27, 523–531. [Google Scholar] [CrossRef]
- Maistrello, L.; Henderson, G.; Laine, R.A. Efficacy of vetiver oil and nootkatone as soil barriers against Formosan subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2001, 94, 1532–1537. [Google Scholar] [CrossRef]
- Alifano, P.; Del Giudice, L.; Talà, A.; De Stefano, M.; Maffei, M.E. Microbes at work in perfumery: The microbial community of vetiver root and its involvement in essential oil biogenesis. Flavour Fragr. J. 2010, 25, 121–122. [Google Scholar] [CrossRef]
- Del Giudice, L.; Massardo, D.R.; Pontieri, P.; Bertea, C.M.; Mombello, D.; Carata, E.; Tredici, S.M.; Talà, A.; Mucciarelli, M.; Ivanova Groudeva, V. The microbial community of Vetiver root and its involvement into essential oil biogenesis. Environ. Microbiol. 2008, 10, 2824–2841. [Google Scholar] [CrossRef]
- Rering, C.; Beck, J.J.; Hall, G.W.; McCartney, M.M.; Vannette, R.L. Nectar-inhabiting microorganisms influence nectar volatile composition and attractiveness to a generalist pollinator. New Phytol. 2017, 3, 750–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schildknecht, H.; Koob, K. Myrmicacin, the first insect herbicide. Angew. Chem. Int. Ed. Engl. 1971, 10, 124–125. [Google Scholar] [CrossRef] [PubMed]
- Iwanami, Y.; Iwadare, T. Myrmic Acids: A Group of New Inhibitors Analogous to Myrmicacin (β-Hydroxydecanoic Acid). Bot. Gaz. 1979, 140, 1–4. [Google Scholar] [CrossRef]
- Sjögren, J.; Magnusson, J.; Broberg, A.; Schnürer, J.; Kenne, L. Antifungal 3-Hydroxy Fatty Acids from Lactobacillus plantarum MiLAB 14. Appl. Environ. Microbiol. 2003, 69, 7554–7557. [Google Scholar] [CrossRef] [Green Version]
- Barke, J.; Seipke, R.F.; Yu, D.W.; Hutchings, M.I. A mutualistic microbiome: How do fungus-growing ants select their antibiotic-producing bacteria? Commun. Integr. Biol. 2011, 4, 41–43. [Google Scholar] [CrossRef]
- Dicke, M. Plant phenotypic plasticity in the phytobiome: A volatile issue. Curr. Opin. Plant Biol. 2016, 32, 17–23. [Google Scholar] [CrossRef]
- Van de Mortel, J.E.; de Vos, R.C.H.; Dekkers, E.; Pineda, A.; Guillod, L.; Bouwmeester, K.; van Loon, J.J.A.; Dicke, M.; Raaijmakers, J.M. Metabolic and Transcriptomic Changes Induced in Arabidopsis by the Rhizobacterium Pseudomonas fluorescens SS101. Plant Physiol. 2012, 160, 2173–2188. [Google Scholar] [CrossRef] [Green Version]
- Pineda, A.; Soler, R.; Weldegergis, B.T.; Shimwela, M.M.; Van Loon, J.J.A.; Dicke, M. Non-pathogenic rhizobacteria interfere with the attraction of parasitoids to aphid-induced plant volatiles via jasmonic acid signalling. Plant Cell Environ. 2013, 36, 393–404. [Google Scholar] [CrossRef]
- Soler, R.; Badenes-Pérez, F.R.; Broekgaarden, C.; Zheng, S.J.; David, A.; Boland, W.; Dicke, M. Plant-mediated facilitation between a leaf-feeding and a phloem-feeding insect in a brassicaceous plant: From insect performance to gene transcription. Funct. Ecol. 2012, 26, 156–166. [Google Scholar] [CrossRef]
- Cusumano, A.; Zhu, F.; Volkoff, A.N.; Verbaarschot, P.; Bloem, J.; Vogel, H.; Dicke, M.; Poelman, E.H. Parasitic wasp-associated symbiont affects plant-mediated species interactions between herbivores. Ecol. Lett. 2018, 21, 957–967. [Google Scholar] [CrossRef] [Green Version]
- Oliver, K.M.; Russell, J.A.; Moran, N.A.; Hunter, M.S. Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA 2003, 100, 1803–1807. [Google Scholar] [CrossRef] [Green Version]
- Frago, E.; Mala, M.; Weldegergis, B.T.; Yang, C.; McLean, A.; Godfray, H.C.J.; Gols, R.; Dicke, M. Symbionts protect aphids from parasitic wasps by attenuating herbivore-induced plant volatiles. Nat. Commun. 2017, 8, 1860. [Google Scholar] [CrossRef] [Green Version]
- Kolesinska, B.; Fraczyk, J.; Binczarski, M.; Modelska, M.; Berlowska, J.; Dziugan, P.; Antolak, H.; Kaminski, Z.J.; Witonska, I.A.; Kregiel, D. Butanol Synthesis Routes for Biofuel Production: Trends and Perspectives. Materials 2019, 12, 350. [Google Scholar] [CrossRef] [Green Version]
- Bowman, K.S.; Rainey, F.A.; Moe, W.M. Production of hydrogen by Clostridium species in the presence of chlorinated solvents. FEMS Microbiol. Lett. 2009, 290, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, F. Über die bedingungen der fuselölbildung und über ihren zusammenhang mit dem eiweissaufbau der hefe. Ber. Dtsch. Chem. Ges. 1907, 40, 1027–1047. [Google Scholar] [CrossRef] [Green Version]
- Vogt, M.; Brüsseler, C.; van Ooyen, J.; Bott, M.; Marienhagen, J. Production of 2-methyl-1-butanol and 3-methyl-1-butanol in engineered Corynebacterium glutamicum. Metab. Eng. 2016, 38, 436–445. [Google Scholar] [CrossRef]
- Hasegawa, M.; Nishiza, A.W.A.; Tsuji, K.; Kimura, S.; Harada, K.I. Volatile Organic Compounds Derived from 2-Keto-Acid Decarboxylase in Microcystis aeruginosa. Microbes Environ. 2012, 27, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Connor, M.R.; Cann, A.F.; Liao, J.C. 3-Methyl-1-butanol production in Escherichia coli: Random mutagenesis and two-phase fermentation. Appl. Microbiol. Biotechnol. 2010, 86, 1155–1164. [Google Scholar] [CrossRef] [Green Version]
- Sato, D.; Nozaki, T. Methionine gamma-lyase: The unique reaction mechanism, physiological roles, and therapeutic applications against infectious diseases and cancers. IUBMB Life 2009, 61, 1019–1028. [Google Scholar] [CrossRef]
- Kadota, H.; Ishida, Y. Production of volatile sulfur compounds by microorganisms. Annu. Rev. Microbiol. 1972, 26, 127–138. [Google Scholar] [CrossRef]
- Nakano, Y.; Yoshimura, M.; Koga, T. Correlation between oral malodor and periodontal bacteria. Microbes Infect. 2002, 4, 679–683. [Google Scholar] [CrossRef]
- Landaud, S.; Helinck, S.; Bonnarme, P. Formation of volatile sulfur compounds and metabolism of methionine and other sulfur compounds in fermented food. Appl. Microbiol. Biotechnol. 2008, 77, 1191–1205. [Google Scholar] [CrossRef] [PubMed]
- Mafra-Neto, A.; Fettig, C.J.; Munson, S.; Stelinski, L.L. Use of repellents formulated in specialized pheromone and lure application technology for effective insect pest management. In Insect Repellents Handbook, 2nd ed.; Debboun, M., Francis, S.P., Strickman, D.A., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2014; Chapter 16; pp. 291–314. ISBN 9781466553583 1466553588; 9781466553552 1466553553. [Google Scholar]
- Tyc, O.; Song, C.; Dickschat, J.S.; Vos, M.; Garbeva, P. The Ecological Role of Volatile and Soluble Secondary Metabolites Produced by Soil Bacteria. Trends Microbiol. 2017, 25, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Cheeseman, G.W.H. Recent advances in pyrazine chemistry. In Advances in Heterocyclic Chemistry; Katritzky, A.R., Boulton, A.J., Eds.; Academic Press Inc.: London, UK, 1972; pp. 99–209. ISBN 978-0120207787. [Google Scholar]
- Kurniadi, T.; Belrhild, R.; Berger, R.F.; Juillerat, M.A.; Fay, L.B. Preparation of Pyrazines. European Patent 1,357,193, 29 October 2003. [Google Scholar]
- Moser, B.R. Review of cytotoxic cephalostatins and ritterazines: Isolation and synthesis. J. Nat. Prod. Rep. 2008, 71, 487–491. [Google Scholar] [CrossRef]
- Prochaska, H.J.; Yeh, Y.; Baron, P.; Polsky, B. Oltipraz (OLT), a novel inhibitor of HIV-1 replication. J. Cell. Biochem. 1995, 59, 117–125. [Google Scholar] [CrossRef]
- Robacker, D.C.; Lauzon, C.R. Purine metabolizing capability of Enterobacter agglomerans affects volatiles production and attractiveness to Mexican fruit fly. J. Chem. Ecol. 2002, 28, 1549–1562. [Google Scholar] [CrossRef]
- Deetae, P.; Bonnarme, P.; Spinnler, H.E.; Henlick, H. Production of volatile aroma compounds by bacterial strains isolated from different surface-ripened french cheeses. Appl. Microbiol. Biotechnol. 2007, 76, 1161–1171. [Google Scholar] [CrossRef]
- Leejeerajumnean, A.; Duckham, S.C.; Owens, J.D.; Ames, J.M. Volatile compounds in Bacillus-fermented soyabeans. J. Sci. Food Agric. 2001, 81, 525–529. [Google Scholar] [CrossRef]
- Rajini, K.S.; Aparna, P.; Sasikala, C.H.; Ramana, C.V. Microbial metabolism of pyrazines. Crit. Rev. Microbiol. 2011, 37, 99–112. [Google Scholar] [CrossRef]
- Beck, H.C.; Hansen, A.M.; Lauritsen, F.R. Novel pyrazine metabolites found in polymyxin biosynthesis by Paenibacillus polymyxa. FEMS Microbiol. Lett. 2003, 220, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Micetich, R.G.; Mac Donald, J.C. Biosynthesis of neoaspergillic acid and neohydroxyaspergillic acids. J. Biol. Chem. 1965, 240, 1692–1695. [Google Scholar] [PubMed]
- Mac Donald, J.C. Toxicity, analyses and production of aspergillic acid and its analogues. Can. J. Biochem. 1973, 51, 1311–1315. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.D.; Kwak, M.K.; Sim, SG. Inhibition of cytochrome P450 2E1 expression by 2-(allylthio) pyrazine, a potential chemoprotective agent: Hepatoprotective effects. Biochem. Pharm. 1997, 53, 261–269. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Houston, W.M. Production of blue-fluorescent pyrazines by Aspergillus parasiticus. J. Food Sci. 1982, 47, 779–782. [Google Scholar] [CrossRef]
- Uffen, R.L.; Canale-Parola, E. Synthesis of pulcherriminic acid by Bacillus subtilis. J. Bacteriol. 1972, 111, 86–93. [Google Scholar] [PubMed]
- Cheng, T.B.; Reineccius, G.A.; Bjorklund, J.A.; Leete, E. Biosynthesis of 2-methoxy-3-isopropylpyrazine in Pseudomonas Perolens. J. Agric. Food Chem. 1991, 39, 1009–1101. [Google Scholar] [CrossRef]
- Gallois, G.; Grimont, P.A.D. Pyrazines responsible for the potatolike odour produced by some Serratia and Cedecea strains. Appl. Environ. Microbiol. 1985, 50, 1048–1051. [Google Scholar]
- Mattey, M.; Harle, E.M. Aerobic metabolism of pyrazine compounds by a Pseudomonas species. Biochem. Soc. Trans. 1976, 4, 492–494. [Google Scholar] [CrossRef] [Green Version]
- Rajini, K.S.; Sasikala, C.; Ramana, C.V. Reductive degradation of pyrazine 2-carboxylate by a newly isolated Stenotrophomonas sp. HCU1. Biodegradation 2010, 21, 801–813. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, J.Y.; Bi, L.J.; Zhou, Y.F.; Zhang, Z.P.; Zhang, C.G.; Zhang, Y.; Zhang, E.X. Characterization of Mycobacterium tuberculosis nicotinamidase/pyrazinamidase. FEBS J. 2008, 275, 753–762. [Google Scholar] [CrossRef]
- Zhang, Y.; Scorpio, A.; Nikaido, H.; Sun, Z. Role of acid pH and deficient efflux of pyrazinoic acid in unique susceptibility of Mycobacterium tuberculosis to pyrazinamide. J. Bacteriol. 1999, 181, 2044–2049. [Google Scholar] [PubMed]
- Zhang, Y.; Wade, M.M.; Scorpio, A.; Zhang, H.; Sun, J. Mode of action of pyrazinamide: Disruption of Mycobacterium tuberculosis membrane transport and energetics by pyrazinoic acid. J. Antimicrob. Chemother. 2003, 52, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Hillier, S.G.; Lathe, R. Terpenes, hormones and life: Isoprene rule revisited. J. Endocrinol. 2019, 242, 2. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Biosynthesis, accumulation and emission of carotenoids, α-tocopherol, plastoquinone, and isoprene in leaves under high photosynthetic irradiance. Photosynth. Res. 2007, 92, 163. [Google Scholar] [CrossRef]
- Villanueva, L.; Sinninghe Damsté, J.S.; Schouten, S. A re-evaluation of the archaeal membrane lipid biosynthetic pathway. Nat. Rev. Microbiol. 2014, 12, 438–448. [Google Scholar] [CrossRef] [Green Version]
- Lehninger, A.L.; Cox, M.M.; Nelson, D.L. Lehninger Principles of Biochemistry, 5th ed.; Freeman, W.H., Ed.; W.H. Freeman and Company: New York, NY, USA, 2008; ISBN 978-1429222631. [Google Scholar]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. Natural products (secondary metabolites). In Biochemistry and Molecular Biology of Plants; Buchanan, B., Gruissem, W., Jones, R., Eds.; American Society of Plant Biologists: Rock-ville, MD, USA, 2000; pp. 1250–1268. [Google Scholar]
- Chappell, J. The genetics and molecular genetics of terpene and sterol origami. Curr. Opin. Plant Biol. 2002, 5, 151–157. [Google Scholar] [CrossRef]
- Mumm, R.; Posthumusm, A.; Dicke, M. Significance of terpenoids in induced indirect plant defence against herbivorous arthropods. Plant Cell Environ. 2008, 31, 575–585. [Google Scholar] [CrossRef]
- Eaton, R.W.; Sandusky, P. Biotransformations of (+/−)-geosmin by terpene-degrading bacteria. Biodegradation 2010, 21, 71. [Google Scholar] [CrossRef]
- Morrone, D.; Chambers, J.; Lowry, L.; Kim, G.; Anterola, A.; Bender, K.; Peters, R.J. Gibberellin biosynthesis in bacteria: Separate ent-copalyl diphosphate and ent-kaurene synthases in Bradyrhizobium japonicum. FEBS Lett. 2009, 583, 475–480. [Google Scholar] [CrossRef] [Green Version]
- Lange, B.M.; Rujan, T.; Martin, W.; Croteau, R. Isoprenoid biosynthesis: The evolution of two ancient and distinct pathways across genomes. Proc. Natl. Acad. Sci. USA 2000, 97, 13172–13177. [Google Scholar] [CrossRef] [Green Version]
- Lombard, J.; Moreira, D. Origins and Early Evolution of the Mevalonate Pathway of Isoprenoid Biosynthesis in the Three Domains of Life. Mol. Biol. Evol. 2011, 28, 87–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffery, C.J. Moonlighting proteins: Old proteins learning new tricks. Trends Genet. 2003, 19, 415–417. [Google Scholar] [CrossRef]
- Zhao, B.; Lei, L.; Vassylyev, D.G.; Lin, X.; Cane, D.E.; Kelly, S.L.; Yuan, H.; Lamb, D.C.; Waterman, M.R. Crystal structure of albaflavenone monooxygenase containing a moonlighting terpene synthase active site. J. Biol. Chem. 2009, 284, 36711–36719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miziorko, H.M. Enzymes of the mevalonate pathway of isoprenoid biosynthesis. Arch. Biochem. Biophys. 2011, 505, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Voynova, N.E.; Rios, S.E.; Miziorko, H.M. Staphylococcus aureus mevalonate kinase: Isolation and characterization of an enzyme of the isoprenoid biosynthetic pathway. J. Bacteriol. 2004, 186, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Wilding, E.I.; Brown, J.R.; Brayant, A.P.; Chalker, A.F.; Holmes, D.J.; Ingrahm, K.A.; Iordanescu, S.; So, C.Y.; Rosenberg, M.; Gwynn, M.N. Identification, Evolution, and Essentiality of the Mevalonate Pathway for Isopentenyl Diphosphate Biosynthesis in Gram-Positive Cocci. J. Bacteriol. 2000, 182, 4319–4327. [Google Scholar] [CrossRef] [Green Version]
- Rodrìguez-Concepcion, M.; Boronat, A. Elucidation of the Methylerythritol Phosphate Pathway for Isoprenoid Biosynthesis in Bacteria and Plastids. A Metabolic Milestone Achieved through Genomics. Plant Physiol. 2002, 130, 1079–1089. [Google Scholar] [CrossRef] [Green Version]
- Brinkman, F.S.; Blanchard, J.L.; Cherkasov, A.; Av-Gay, Y.; Brunham, R.C.; Fernandez, R.C.; Finlay, B.B.; Otto, S.P.; Ouellette, B.F.; Keeling, P.J.; et al. Evidence that plant-like genes in Chlamydia species reflect an ancestral relationship between Chlamydiaceae, cyanobacteria, and the chloroplast. Genome Res. 2002, 12, 1159–1167. [Google Scholar] [CrossRef] [Green Version]
- Rohdich, F.; Kis, K.; Bacher, A.; Eisenreich, W. The non-mevalonate pathway of isoprenoids: Genes, enzymes and intermediates. Curr. Opin. Chem. Biol. 2001, 5, 535–540. [Google Scholar] [CrossRef]
- Sprenger, G.A.; Schorken, U.; Wiegert, T.; Grolle, S.; de Graaf, A.A.; Taylor, S.V.; Begley, T.P.; Bringer-Meyer, S.; Sahm, H. Identification of a thiamin-dependent synthase in Escherichia coli required for the formation of the 1-deoxy-D-xylulose 5-phosphate precursor to isoprenoids, thiamin, and pyridoxol. Proc. Natl. Acad. Sci. USA 1997, 94, 12857–12862. [Google Scholar] [CrossRef] [Green Version]
- Lois, L.M.; Campos, N.; Rosa Putra, S.; Danielsen, K.; Rohmer, M.; Boronat, A. Cloning and characterization of a gene from Escherichia coli encoding a transketolase-like enzyme that catalyzes the synthesis of D-1-deoxyxylulose 5-phosphate, a common precursor for isoprenoid, thia- min, and pyridoxol biosynthesis. Proc. Natl. Acad. Sci. USA 1998, 95, 2105–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzuyama, T.; Takahashi, S.; Watanabe, H.; Seto, H. Direct formation of 2-C-methyl-D-erythritol 4-phosphate from 1-deoxy-D-xylulose 5-phosphate by 1-deoxy-D-xylulose 5-phosphate reductoisomerase, a new enzyme in the non-mevalonate pathway to isopentenyl diphosphate. Tetrahedron Lett. 1998, 39, 4509–4512. [Google Scholar] [CrossRef]
- Takahashi, S.; Kuzuyama, T.; Watanabe, H.; Seto, H. A 1-deoxy-D- xylulose 5-phosphate reductoisomerase catalyzing the formation of 2-C- methyl-D-erythritol 4-phosphate in an alternative nonmevalonate pathway for terpenoid biosynthesis. Proc. Natl. Acad. Sci. USA 1998, 95, 9879–9884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohdich, F.; Wungsintaweekul, J.; Fellermeier, M.; Sagner, S.; Herz, S.; Kis, K.; Eisenreich, W.; Bacher, A.; Zenk, M.H. Cytidine 5′-triphosphate- dependent biosynthesis of isoprenoids: YgbP protein of Escherichia coli catalyzes the formation of 4-diphosphocytidyl-2-C-methylerythritol. Proc. Natl. Acad. Sci. USA 1999, 96, 11758–11763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luttgen, H.; Rohdich, F.; Herz, S.; Wungsintaweekul, J.; Hecht, S.; Schuhr, C.A.; Fellermeier, M.; Sagner, S.; Zenk, M.H.; Bacher, A.; et al. Biosynthesis of terpenoids: YchB protein of Escherichia coli phosphorylates the 2-hydroxy group of 4-diphosphocytidyl-2-C-methyl-D-erythritol. Proc. Natl. Acad. Sci. USA 2000, 97, 1062–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herz, S.; Wungsintaweekul, J.; Schuhr, C.A.; Hecht, S.; Luttgen, H.; Sagner, S.; Fellermeier, M.; Eisenreich, W.; Zenk, M.H.; Bacher, A.; et al. Biosynthesis of terpenoids: YgbB protein converts 4-diphosphocytidyl- 2C-methyl-D-erythritol 2-phosphate to 2C-methyl-D-erythritol 2,4- cyclodiphosphate. Proc. Natl. Acad. Sci. USA 2000, 97, 2486–2490. [Google Scholar] [CrossRef] [Green Version]
- Rohdich, F.; Hecht, S.; Gartner, K.; Adam, P.; Krieger, C.; Amslinger, S.; Arigoni, D.; Bacher, A.; Eisenreich, W. Studies on the nonmevalonate terpene biosynthetic pathway: Metabolic role of IspH (LytB) protein. Proc. Natl. Acad. Sci. USA 2002, 99, 1158–1163. [Google Scholar] [CrossRef] [Green Version]
- Hahn, F.M.; Hurlburt, A.P.; Poulter, C.D. Escherichia coli open reading frame 696 is idi, a nonessential gene encoding isopentenyl diphosphate isomerase. J. Bacteriol. 1999, 181, 4499–4504. [Google Scholar]
- Nakano, C.; Kim, H.K.; Ohnishi, Y. Identification of the first bacterial monoterpene cyclase, a 1,8-cineole synthase, that catalyzes the direct conversion of geranyl diphosphate. ChemBioChem 2011, 5, 1988–1991. [Google Scholar] [CrossRef]
- Köksal, M.; Chou, W.K.; Cane, D.E.; Christianson, D.W. Unexpected reactivity of 2-fluorolinalyl diphosphate in the active site of crystalline 2-methylisoborneol synthase. Biochemistry 2013, 6, 5247–5255. [Google Scholar] [CrossRef] [Green Version]
- Nakano, C.; Kim, H.K.; Ohnishi, Y. Identification and characterization of the linalool/nerolidol synthase from Streptomyces clavuligerus. ChemBioChem 2011, 4, 2403–2407. [Google Scholar] [CrossRef] [PubMed]
- Dickschat, J.S.; Pahirulzaman, K.A.; Rabe, P.; Klapschinski, T.A. An improved technique for the rapid chemical characterisation of bacterial terpene cyclases. ChemBioChem 2014, 14, 810–814. [Google Scholar] [CrossRef]
- Brown, W.H.; Foote, C.S.; Iverson, B.L.; Anslyn, E.V. Organic Chemistry, 5th ed.; Harcourt College Publishers: Austin, TX, USA, 2007; ISBN 978-0981519432. [Google Scholar]
- Koe, B.K.; Sobin, B.A.; Celmer, W.D. PA 132, a new antibiotic. I. Isolation and chemical properties. Antibiot. Annu. 1956, 672, 5. [Google Scholar]
- Cane, D.E.; Watt, R.M. Expression and mechanistic analysis of a germacradienol synthase from Streptomyces coelicolor implicated in geosmin biosynthesis. Proc. Natl. Acad. Sci. USA 2003, 18, 1547–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagrain, B.; Martens, J.; Roelants, I.; Wenseleers, T. Method for Controlling Ant Colonies. Worldwide Application WO2015081390A3, 11 June 2015. [Google Scholar]
- Kim, S.Y.; Zhao, P.; Igarashi, M.; Sawa, R.; Tomita, T.; Nishiyama, M.; Kuzuyama, T. Cloning and heterologous expression of the cyclooctatin biosynthetic gene cluster afford a diterpene cyclase and two p450 hydroxylases. Chem. Biol. 2009, 16, 736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorner, C.; Hauslein, I.; Schrepfer, P.; Eisenreich, W.; Bruck, T. Targeted Engineering of Cyclooctat-9-en-7-ol Synthase: A Stereospecific Access to Two New Non-natural Fusicoccane-Type Diterpenes. ChemCatChem 2013, 5, 3289. [Google Scholar] [CrossRef]
- Dairi, T.; Hamano, Y.; Kuzuyama, T.; Itoh, N.; Furihata, K.; Seto, H. Eubacterial diterpene cyclase genes essential for production of the isoprenoid antibiotic terpentecin. J. Bacteriol. 2001, 183, 6085. [Google Scholar] [CrossRef] [Green Version]
- Hamano, Y.; Kuzuyama, T.; Itoh, N.; Furihata, K.; Seto, H.; Dairi, T. Functional analysis of eubacterial diterpene cyclases responsible for biosynthesis of a diterpene antibiotic, terpentecin. J. Biol. Chem. 2002, 277, 37098. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Lu, X. Microbial Synthesis of Alka(e)nes. Front. Bioeng. Biotechnol. 2013, 1, 10. [Google Scholar] [CrossRef] [Green Version]
- Schirmer, A.; Rude, M.A.; Li, X.Z.; Popova, E.; del Cardayre, S.B. Microbial biosynthesis of alkanes. Science 2010, 329, 559–562. [Google Scholar] [CrossRef]
- Li, N.; Chang, W.C.; Warui, D.M.; Booker, S.J.; Krebs, C.; Bollinger, J.M. Evidence for only oxygenative cleavage of aldehydes to alk(a/e)nes and formate by cyanobacterial aldehyde decarbonylases. Biochemistry 2012, 51, 7908–7916. [Google Scholar] [CrossRef] [PubMed]
- Beller, H.R.; Goh, E.B.; Keasling, J.D. Genes involved in long- chain alkene biosynthesis in Micrococcus luteus. Appl. Environ. Microbiol. 2010, 76, 1212–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rude, M.A.; Baron, T.S.; Brubaker, S.; Alibhai, M.; Del Cardayre, S.B.; Schirmer, A. Terminal olefin (1-alkene) biosynthesis by a novel P450 fatty acid decarboxylase from Jeotgalicoccus species. Appl. Environ. Microbiol. 2011, 77, 1718–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendez-Perez, D.; Begemann, M.B.; Pfleger, B.F. Modular synthase-encoding gene involved in alpha-olefin biosynthesis in Synechococcus sp strain PCC 7002. Appl. Environ. Microbiol. 2011, 77, 4264–4267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhtar, M.K.; Turner, N.J.; Jones, P.R. Carboxylic acid reductase is a versatile enzyme for the conversion of fatty acids into fuels and chemical commodities. Proc. Natl. Acad. Sci. USA 2013, 110, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, T.P.; Middelhaufe, S.; Moore, K.; Edner, C.; Kolak, D.M.; Taylor, G.N. Synthe- sis of customized petroleum-replica fuel molecules by targeted modification of free fatty acid pools in Escherichia coli. Proc. Natl. Acad. Sci. USA 2013, 110, 7636–7641. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Tittiger, C.; Wicker-Thomas, C.; Le Goff, G.; Young, S.; Wajnberg, E.; Fricaux, T.; Taquet, N.; Blomquist, G.J.; Feyereisen, R. An insect-specific P450 oxidative decarbonylase for cuticular hydrocarbon biosynthesis. Proc. Natl. Acad. Sci. USA 2012, 109, 14858–14863. [Google Scholar] [CrossRef] [Green Version]
- Garbeva, P.; Hordijk, C.; Gerards, S.; de Boer, W. Volatile-mediated interactions between phylogenetically different soil bacteria. Front. Microbiol. 2014, 5, 289. [Google Scholar] [CrossRef] [Green Version]
- Piechulla, B.; Lemfack, M.C.; Kai, M. Effects of discrete bioactive microbial volatiles on plants and fungi. Plant Cell Environ. 2017, 40, 2042–2067. [Google Scholar] [CrossRef]
- Schulz-Bohm, K.; Zweers, H.; de Boer, W.; Garbeva, P. A fragrant neighborhood: Volatile mediated bacterial interactions in soil. Front. Microbiol. 2015, 6, 1212. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, R.; Cordovez, V.; de Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyc, O.; Wolf, AB.; Garbeva, P. The effect of phylogenetically different bacteria on the fitness of Pseudomonas fluorescens in sand microcosms. PLoS ONE 2015, 10, e0119838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyc, O.; de Jager, V.C.L.; van den Berg, M.; Gerards, S.; Janssens, T.K.S.; Zaagman, N.; Kai, M.; Svatos, A.; Zweers, H.; Hordijk, C.; et al. Exploring bacterial interspecific interactions for discovery of novel antimicrobial compounds. Microb. Biotechnol. 2017, 10, 910–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Cordovez, V.; Etalo, D.W.; van der Voort, M.; Raaijmakers, J.M. Role of the GacS sensor kinase in the regulation of volatile production by plant growth-promoting Pseudomonas fluorescens SBW25. Front. Plant Sci. 2016, 7, 1706. [Google Scholar] [CrossRef] [PubMed]
- Lapouge, K.; Schubert, M.; Allain, F.H.; Haas, D. Gac/Rsm signal transduction pathway of gamma-proteobacteria: From RNA recognition to regulation of social behaviour. Mol. Microbiol. 2008, 67, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Heeb, S.; Haas, D. Regulatory roles of the GacS/GacA two-component system in plant-associated and other gram-negative bacteria. Mol. Plant Microb. Interact. 2001, 14, 1351–1363. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Bacteria and insect communication. In S. gregaria (A), the bacterium P. agglomerans produces guaiacol, which acts as an aggregation signal. In D. valens (B), bacterial community convert conifer monoterpenoids in verbenone, a multifunctional pheromone. In B. germanica (C), the aggregation is stimulated by succinic acid and other acids produced by gut bacteria. In A. gambiae (D), pyrazines, carboxylic acids, and alcohols stimulate oviposition. In P. regina (E), bacterial volatile organic compounds (BVOCs) stimulate attraction between individuals. In eusocial insects (F) (Hymenoptera), social and behavioral skills are managed by pyrazines, cuticular hydrocarbons, and other semiochemicals.
Figure 1.
Bacteria and insect communication. In S. gregaria (A), the bacterium P. agglomerans produces guaiacol, which acts as an aggregation signal. In D. valens (B), bacterial community convert conifer monoterpenoids in verbenone, a multifunctional pheromone. In B. germanica (C), the aggregation is stimulated by succinic acid and other acids produced by gut bacteria. In A. gambiae (D), pyrazines, carboxylic acids, and alcohols stimulate oviposition. In P. regina (E), bacterial volatile organic compounds (BVOCs) stimulate attraction between individuals. In eusocial insects (F) (Hymenoptera), social and behavioral skills are managed by pyrazines, cuticular hydrocarbons, and other semiochemicals.

Figure 2.
Aphids mutualism and predation. In aphids, S. xylosus and S. sciuri produces BVOCs that acts as an attractant for ants. Mutualistic ants manage and protect aphid nests. On the other hand, S. sciuri and A. calcoaceticus produce some BVOCs that help E. blateatus larvae to find the aphids (the prey).
Figure 2.
Aphids mutualism and predation. In aphids, S. xylosus and S. sciuri produces BVOCs that acts as an attractant for ants. Mutualistic ants manage and protect aphid nests. On the other hand, S. sciuri and A. calcoaceticus produce some BVOCs that help E. blateatus larvae to find the aphids (the prey).

Figure 3.
Mirmecophilus butterfly. P. alcon (Maculinea) is a lepidoptera that is adopted by ants (M. scabrinodis) in, intermediate larvae (IL) stage when the larvae fall on soil (after the early phytophagous phase). The attraction mechanism and chemical camouflage strategy is related to gut microbiota, which produces BVOCs. The genera Staphylococcus and Serratia shows a consistent distribution with camouflage skills. PICRUST prediction of genetic content shows as digestion of carbohydrate and proteins are consistent with the development of larvae and validate the system.
Figure 3.
Mirmecophilus butterfly. P. alcon (Maculinea) is a lepidoptera that is adopted by ants (M. scabrinodis) in, intermediate larvae (IL) stage when the larvae fall on soil (after the early phytophagous phase). The attraction mechanism and chemical camouflage strategy is related to gut microbiota, which produces BVOCs. The genera Staphylococcus and Serratia shows a consistent distribution with camouflage skills. PICRUST prediction of genetic content shows as digestion of carbohydrate and proteins are consistent with the development of larvae and validate the system.

Figure 4.
Pyrazines biosynthesis. Generally, experimental data show a correlation between amino acids metabolisms and pyrazine production in microorganisms. Yeast species, if cultured in isoleucine or leucine-rich medium, produce pyrazines used in an industrial process (A) Aspergillus produces aspergillic acid and similar molecules, Candida pulcherrima produces pulcherriminc acid. A similar effect (B) is observed in Pseudomonas perolens, adding valine and glycine in the culture broth. In this last case, there was a proposed pathway. Finally, adding radiolabeled precursors, a pathway was individuated in Serratia marcescens B2 that start from threonine (C).
Figure 4.
Pyrazines biosynthesis. Generally, experimental data show a correlation between amino acids metabolisms and pyrazine production in microorganisms. Yeast species, if cultured in isoleucine or leucine-rich medium, produce pyrazines used in an industrial process (A) Aspergillus produces aspergillic acid and similar molecules, Candida pulcherrima produces pulcherriminc acid. A similar effect (B) is observed in Pseudomonas perolens, adding valine and glycine in the culture broth. In this last case, there was a proposed pathway. Finally, adding radiolabeled precursors, a pathway was individuated in Serratia marcescens B2 that start from threonine (C).

Figure 5.
Terpene and terpenoids biosynthesis. The precursors for biosynthesis of terpenes and terpenoids (isopentyl-pyroP, IPP and dimethylallyl-piroP, DMAPP) are synthetized by two biosynthetic pathways: The mevalonate pathway (MVA, in red) and the methylerythritol phosphate pathway (MEP, in green). As shown in the the square with blue dotted line, MVA was found in eukaryotes and archaea, while MEP was found in bacteria or in eukaryotes equipped with plastid. Horizontal gene exchange has allowed the acquisition of MVA (for the bacteria) or MEP genes (for the eukaryotes). GPS (geranyl-pyroP synthase), FPS (farnesyl-pyroP synthase) and GGPS (geranylgeranyl-pyroP synthase) assemble IPP and DMAPP into linear long-chain precursors used by the biosynthetic enzymes to produce the terpenes-like ring. In the figure, the examples are reported (from precursor to ring cyclization), including monoterpene synthases, diterpene synthases, and sesquiterpene synthases. Abbreviations: MVA: AACT = acetoacetyl-CoA thiolase, HMGS = 3-hydroxy-3-methylglutaryl- CoA synthase, HMGR = 3-hydroxy-3-methylglutaryl- CoA reductase, MVK = mevalonate kinase, PMK = phosphomevalonate kinase, MDC = mevalonate- 5-decarboxylase. Abbreviation of MEP: DXS = 1-deoxy-d-xylulose-5-phosphate synthase, DXP = deoxy-d-xylulose-5-phosphate, DXR = 1-deoxy-d-xylulose-5-reductase; MEP = methyl-erythritol phosphate, CMS = MEP cytidylyltransferase, CDP-MEP = cytidylyl-MEP, MCS = ME-cPP synthase, ME-cPP = methyl-erythritol 2,4-cyclodiphosphate (ME-cPP), HDS = hydroxymethylbutenyl 4-diphosphate synthase, HMBPP = hydroxymethylbutenyl 4-diphosphate, IDS = IPP/DMAPP synthase, IDI1/IDI2 = IPP-DAMPP isomerase.
Figure 5.
Terpene and terpenoids biosynthesis. The precursors for biosynthesis of terpenes and terpenoids (isopentyl-pyroP, IPP and dimethylallyl-piroP, DMAPP) are synthetized by two biosynthetic pathways: The mevalonate pathway (MVA, in red) and the methylerythritol phosphate pathway (MEP, in green). As shown in the the square with blue dotted line, MVA was found in eukaryotes and archaea, while MEP was found in bacteria or in eukaryotes equipped with plastid. Horizontal gene exchange has allowed the acquisition of MVA (for the bacteria) or MEP genes (for the eukaryotes). GPS (geranyl-pyroP synthase), FPS (farnesyl-pyroP synthase) and GGPS (geranylgeranyl-pyroP synthase) assemble IPP and DMAPP into linear long-chain precursors used by the biosynthetic enzymes to produce the terpenes-like ring. In the figure, the examples are reported (from precursor to ring cyclization), including monoterpene synthases, diterpene synthases, and sesquiterpene synthases. Abbreviations: MVA: AACT = acetoacetyl-CoA thiolase, HMGS = 3-hydroxy-3-methylglutaryl- CoA synthase, HMGR = 3-hydroxy-3-methylglutaryl- CoA reductase, MVK = mevalonate kinase, PMK = phosphomevalonate kinase, MDC = mevalonate- 5-decarboxylase. Abbreviation of MEP: DXS = 1-deoxy-d-xylulose-5-phosphate synthase, DXP = deoxy-d-xylulose-5-phosphate, DXR = 1-deoxy-d-xylulose-5-reductase; MEP = methyl-erythritol phosphate, CMS = MEP cytidylyltransferase, CDP-MEP = cytidylyl-MEP, MCS = ME-cPP synthase, ME-cPP = methyl-erythritol 2,4-cyclodiphosphate (ME-cPP), HDS = hydroxymethylbutenyl 4-diphosphate synthase, HMBPP = hydroxymethylbutenyl 4-diphosphate, IDS = IPP/DMAPP synthase, IDI1/IDI2 = IPP-DAMPP isomerase.

Figure 6.
Linear hydrocarbons biosynthesis. Microorganisms produce hydrocarbons through different biosynthetic pathways. The blue arrows indicate bacterial metabolism for a precursor of hydrocarbons (propionic acid). As shown, different pathways produce propionate including fermentation, biotransformation of succinate, and biotransformation of intermediates from amino acid metabolisms. In Z. nevadensis, propionate is produced by bacterial community starting from succinate and is then assembled in final cuticular hydrocarbons by the insect. The green arrows show the direct synthesis of alkanes and alkenes in bacteria. There are four pathways: AAR/ADO, Ols, OleABCD, and OleTJE. Abbreviations: SCS = succinyl-CoA synthetase, MUT = methylmalonyl-CoA mutase, MCEE = methylmalonyl-CoA/ethylmalonyl-CoA epimerase, ScpB = methylmalonyl-CoA decarboxylase, PCCA = propionyl-CoA carboxylase alpha chain, EC 2.1.3.1 = methylmalonyl-CoA carboxytransferase, PflD = formate C-acetyltransferase, PorA = pyruvate ferredoxin oxidoreductase alpha subunit, Pta = phosphate acetyltransferase, PduL = phosphate propanoyltransferase, AckA = acetate kinase, TdcD = propionate kinase; Pct = propionate CoA-transferase, AcdA = acetate-CoA ligase (ADP-forming) subunit alpha.
Figure 6.
Linear hydrocarbons biosynthesis. Microorganisms produce hydrocarbons through different biosynthetic pathways. The blue arrows indicate bacterial metabolism for a precursor of hydrocarbons (propionic acid). As shown, different pathways produce propionate including fermentation, biotransformation of succinate, and biotransformation of intermediates from amino acid metabolisms. In Z. nevadensis, propionate is produced by bacterial community starting from succinate and is then assembled in final cuticular hydrocarbons by the insect. The green arrows show the direct synthesis of alkanes and alkenes in bacteria. There are four pathways: AAR/ADO, Ols, OleABCD, and OleTJE. Abbreviations: SCS = succinyl-CoA synthetase, MUT = methylmalonyl-CoA mutase, MCEE = methylmalonyl-CoA/ethylmalonyl-CoA epimerase, ScpB = methylmalonyl-CoA decarboxylase, PCCA = propionyl-CoA carboxylase alpha chain, EC 2.1.3.1 = methylmalonyl-CoA carboxytransferase, PflD = formate C-acetyltransferase, PorA = pyruvate ferredoxin oxidoreductase alpha subunit, Pta = phosphate acetyltransferase, PduL = phosphate propanoyltransferase, AckA = acetate kinase, TdcD = propionate kinase; Pct = propionate CoA-transferase, AcdA = acetate-CoA ligase (ADP-forming) subunit alpha.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Calcagnile, M.; Tredici, S.M.; Talà, A.; Alifano, P. Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants. Insects 2019, 10, 441. https://doi.org/10.3390/insects10120441
AMA Style
Calcagnile M, Tredici SM, Talà A, Alifano P. Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants. Insects. 2019; 10(12):441. https://doi.org/10.3390/insects10120441
Chicago/Turabian StyleCalcagnile, Matteo, Salvatore Maurizio Tredici, Adelfia Talà, and Pietro Alifano. 2019. "Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants" Insects 10, no. 12: 441. https://doi.org/10.3390/insects10120441
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.