Modern Techniques in Colorado Potato Beetle (Leptinotarsa decemlineata Say) Control and Resistance Management: History Review and Future Perspectives



Abstract
:Simple Summary
Abstract
1. Introduction
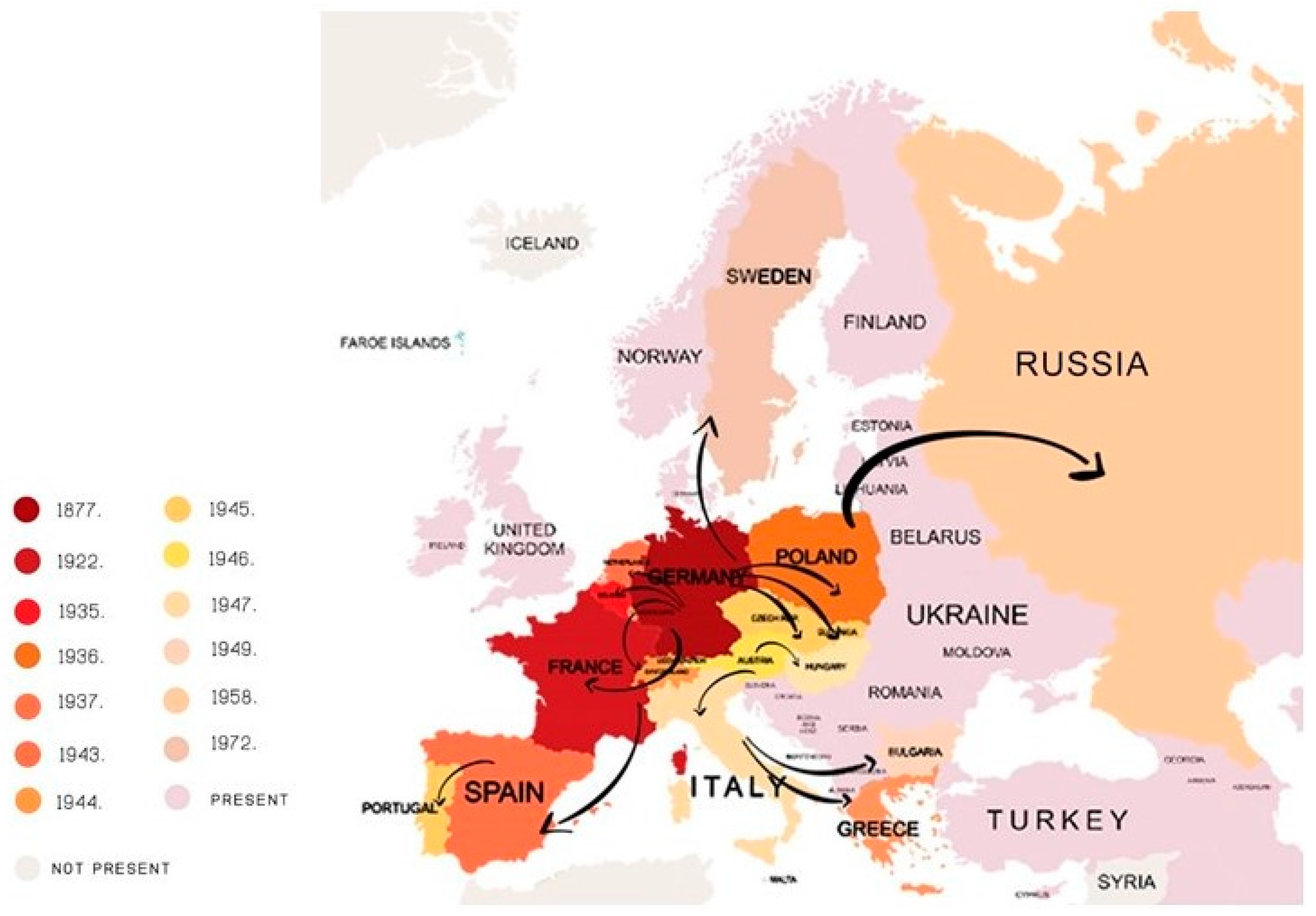
Colorado Potato Beetle—A Global Pest of Potato Production
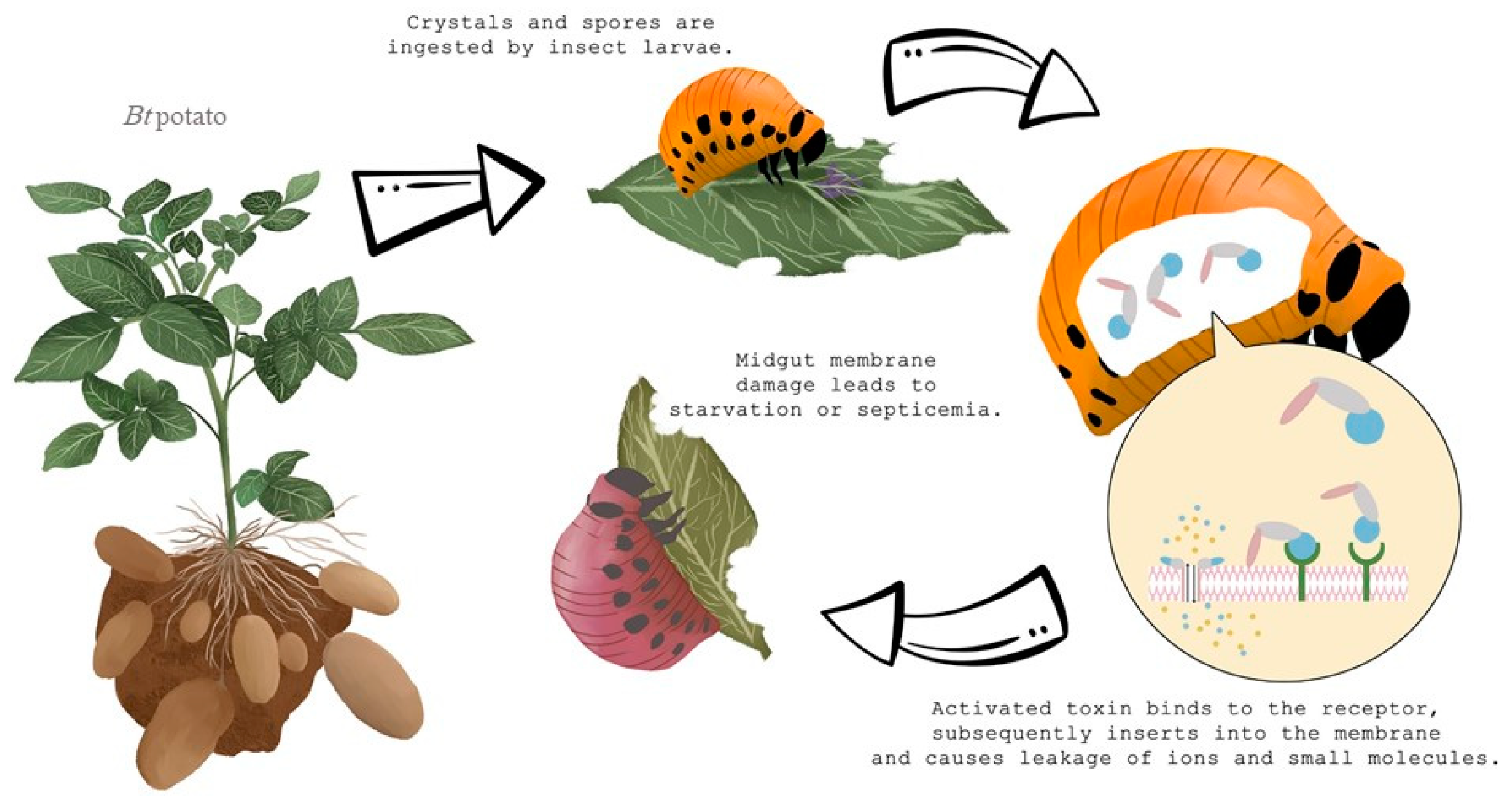
2. Bacillus thuringiensis (Bt) in the Fight to Control Colorado Potato Beetle
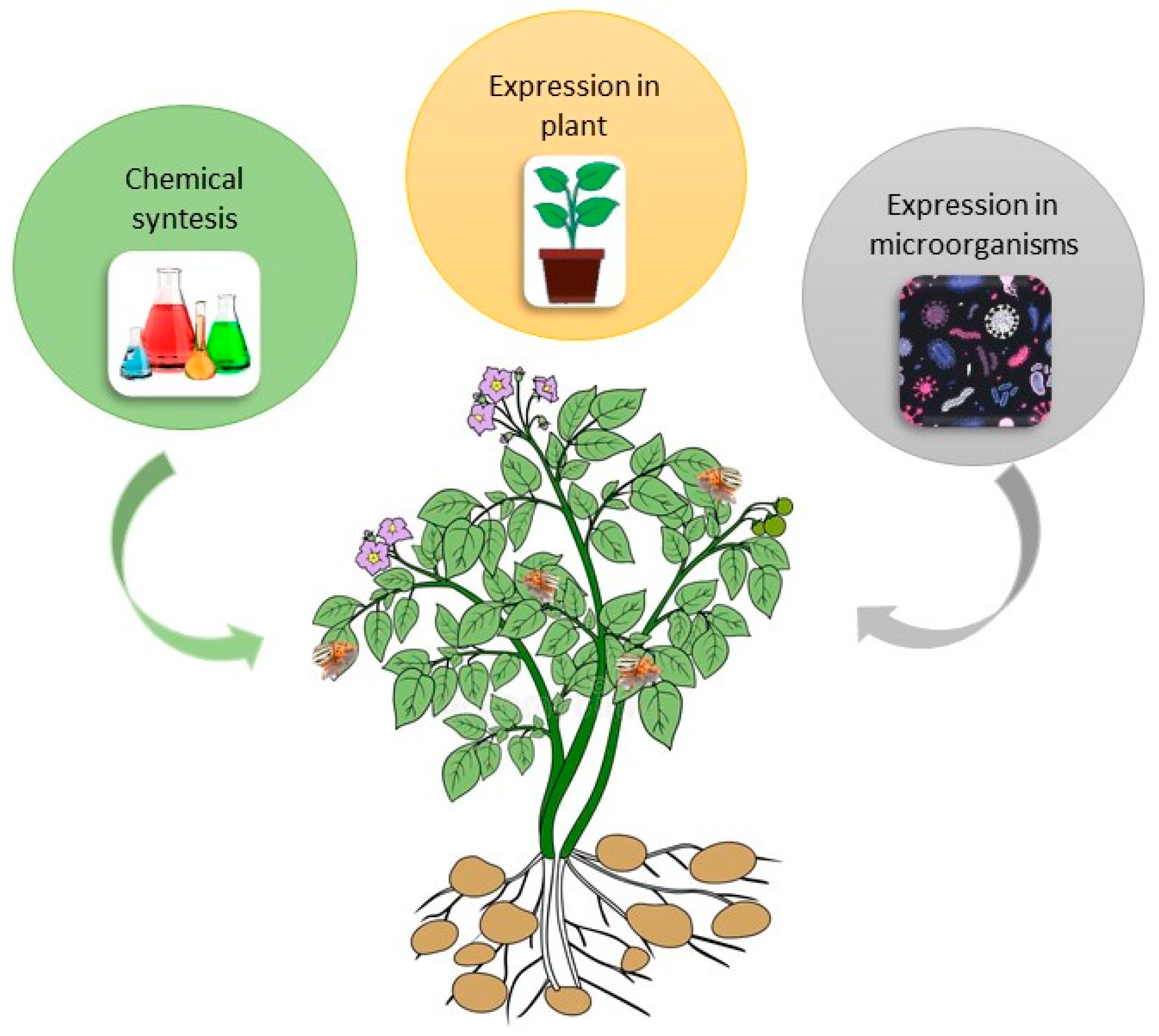
2.1. Bt Potato Development
2.2. Why Bt Potato Did Not Sustain on Market
3. Sources of Host-Plant Resistance
4. New Approaches to Colorado Potato Beetle Management
4.1. RNA Interference (RNAi)
4.2. RNAi in Colorado Potato Beetle Control Management
4.3. RNAi Based Products in Wide Use
5. Genetic Tools in Colorado Potato Beetle Management
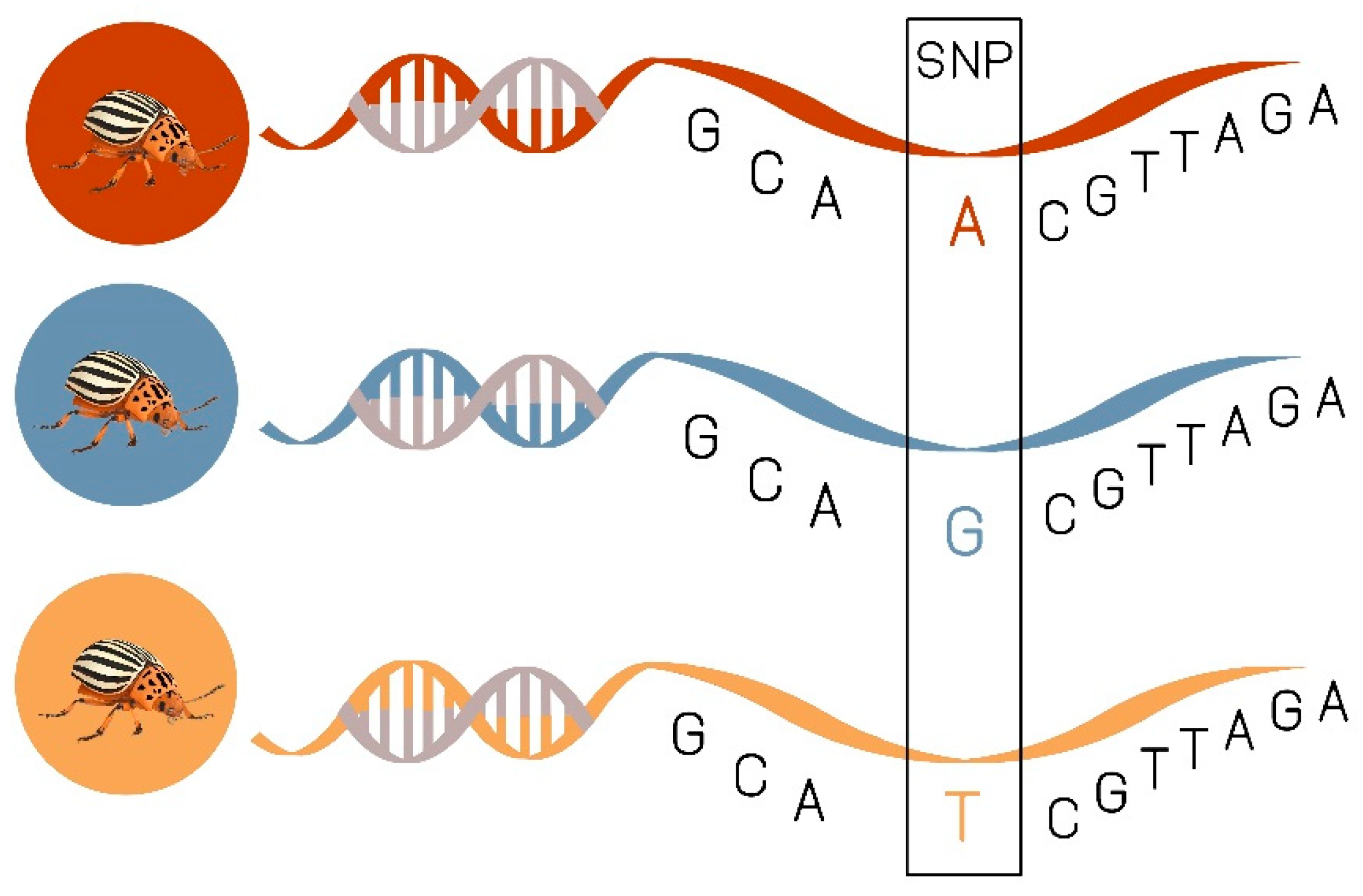
Single Nucleotide Polymorphism (SNPs) as Prospective Tool in CPB Resistance Management
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations FAO STAT. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 31 March 2020).
- James, C. Global Status of Commercialized Biotech/GM Crops; ISAAA: Ithaca, NY, USA, 2011; Volume 44. [Google Scholar]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Radcliffe, E.B.; Lagnaoui, A. Pests and Diseases. In Potato Biology and Biotechnology: Advances and Perspectives, 1st ed.; Vreugdenhil, D., Bradshaw, J., Gebhardt, C., Govers, F., Taylor, M.A., MacKerron, D.K., Ross, H.A., Eds.; Elsevier: Oxford, UK, 2007; pp. 545–554. [Google Scholar]
- Weber, D. Colorado beetle: Pest on the move. Pestic. Outlook 2003, 14, 256–259. [Google Scholar] [CrossRef]
- Alyokhin, A. Colorado potato beetle management on potatoes: Current challenges and future prospects. Fruit Veg. Cereal Sci. Biotechnol. 2009, 3, 10–19. [Google Scholar]
- Riley, C.V. Seventh Annual Report on the Noxious, Beneficial, and Other Insects of the State of Missouri, 1st ed.; Regan & Carter: Jefferson City, MO, USA, 1875; pp. 1–50. [Google Scholar]
- Casagrande, R.A. The Colorado potato beetle: 125 years of mismanagement. Bull. Entomol. Soc. Am. 1987, 33, 142–150. [Google Scholar] [CrossRef]
- Alyokhin, A.; Baker, M.; Mota-Sanchez, D.; Dively, G.; Grafius, E. Colorado potato beetle resistance to insecticides. Am. J. Potato Res. 2008, 85, 395–413. [Google Scholar] [CrossRef]
- Cong, W.A.N.G.; Han, X.U.; Pan, X.B. Management of Colorado potato beetle in invasive frontier areas. J. Integr. Agric. 2020, 19, 360–366. [Google Scholar]
- Ferro, D.N.; Logan, J.A.; Voss, R.H.; Elkinton, J.S. Colorado potato beetle (Coleoptera: Chrysomelidae) temperature-dependent growth and feeding rates. Environ. Entomol. 1985, 14, 343–348. [Google Scholar] [CrossRef]
- Kennedy, G.G. Colorado potato beetle. In Encyclopedia of Insects, 1st ed.; Academic Press: Cambridge, MA, USA, 2009; pp. 212–213. [Google Scholar]
- Alyokhin, A.; Udalov, M.; Benkovskaya, G. The Colorado potato beetle. Insect Pests of Potato. Glob. Perspect. Biol. Manag. 2013, 2, 11. [Google Scholar]
- Maharijaya, A.; Vosman, B. Managing the Colorado potato beetle; the need for resistance breeding. Euphytica 2015, 204, 487–501. [Google Scholar] [CrossRef] [Green Version]
- Kiss, J.; Komaromi, J.; Bayar, K.; Edwards, C.R.; Hatala-Zseller, I. Western corn rootworm (Diabrotica virgifera virgifera LeConte) and the crop rotation systems in Europe. In Western Corn Rootworm: Ecology and Management, 1st ed.; Vidal, S., Kuhlmann, U., Edwards, C.R., Eds.; CAB International: Wallingford, UK, 2005; pp. 189–220. [Google Scholar]
- Grafius, E.J.; Douches, D.S. The present and future role of insect-resistant genetically modified potato cultivars in IPM. In Integration of Insect-Resistant Genetically Modified Crops within IPM Programs, 1st ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 195–221. [Google Scholar]
- Gauthier, N.L.; Hofmaster, R.N.; Semel, M. History of Colorado potato beetle control. Adv. Potato Pest Manag. 1981, 23, 13–33. [Google Scholar]
- Grafius, E. Economic impact of insecticide resistance in the Colorado potato beetle (Coleoptera: Chrysomelidae) on the Michigan potato industry. J. Econ. Entomol. 1997, 90, 1144–1151. [Google Scholar] [CrossRef]
- Stanković, S.; Zabel, A.; Kostic, M.; Manojlovic, B.; Rajkovic, S. Colorado potato beetle [Leptinotarsa decemlineata (Say)] resistance to organophosphates and carbamates in Serbia. J. Pest Sci. 2004, 77, 11–15. [Google Scholar] [CrossRef]
- Sladan, S.; Miroslav, K.; Ivan, S.; Snezana, J.; Petar, K.; Goran, T.; Jevdovic, R. Resistance of Colorado potato beetle (Coleoptera: Chrysomelidae) to neonicotinoids, pyrethroids and nereistoxins in Serbia. Rom. Biotechnol. Lett. 2012, 17, 7599–7609. [Google Scholar]
- Szendrei, Z.; Grafius, E.; Byrne, A.; Ziegler, A. Resistance to neonicotinoid insecticides in field populations of the Colorado potato beetle (Coleoptera: Chrysomelidae). Pest Manag. Sci. 2012, 68, 941–946. [Google Scholar] [CrossRef]
- Scott, I.M.; Tbeetle Leptinotaolman, J.H.; MacArthur, D.C. Insecticide resistance and cross-resistance development in Colorado potato rsa decemlineata Say (Coleoptera: Chrysomelidae) populations in Canada 2008–2011. Pest Manag. Sci. 2015, 71, 712–721. [Google Scholar] [CrossRef]
- Arthropod Pesticide Resistance Database (APRD). Leptinotarsa decemlineata-Shown Resistance to Active Ingredient(s). Available online: https://www.pesticideresistance.org/display.php?page=species&arId=141 (accessed on 26 February 2020).
- Hellmich, R.L.; Albajes, R.; Bergvinson, D.; Prasifka, J.R.; Wang, Z.Y.; Weiss, M.J. The present and future role of insect-resistant genetically modified maize in IPM. In Integration of Insect-Resistant Genetically Modified Crops within IPM Programs, 1st ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 119–158. [Google Scholar]
- Abbas, M.S.T. Genetically engineered (modified) crops (Bacillus thuringiensis crops) and the world controversy on their safety. Egypt. J. Biol. Pest Control 2018, 28, 1–12. [Google Scholar] [CrossRef]
- SPUDsmart. Potato Breeding: A European Approach, Part III. Available online: https://spudsmart.com/potato-breeding-a-european-approach-part-iii/ (accessed on 20 May 2020).
- European Food Safety Authority (EFSA). Available online: http://www.efsa.europa.eu/ (accessed on 30 March 2020).
- Thompson, A.L.; Farnsworth, B.L.; Gudmestad, N.C.; Secor, G.A.; Preston, D.A.; Sowokinos, J.R.; Glynn, M.; Hatterman-Valenti, H. Dakota diamond: An exceptionally high yielding, cold chipping potato cultivar with long-term storage potential. Am. J. Potato Res. 2008, 85, 171. [Google Scholar] [CrossRef]
- Dik, A.; Ceglarska, E.; Ilovai, Z. Sweet pepper: Development in plant pathology. In Integrated Pest and Disease Management in Greenhouse Crops; Springer: Dodrecht, The Netherlands, 2000; pp. 473–485. [Google Scholar]
- Walker, K.; Mendelsohn, M.; Matten, S.; Alphin, M.; Ave, D. The role of microbial Bt products in US crop protection. J. New Seeds 2003, 5, 31–51. [Google Scholar] [CrossRef]
- Perlak, F.J.; Stone, T.B.; Muskopf, Y.M.; Petersen, L.J.; Parker, G.B.; McPherson, S.A.; Wyman, J.; Love, S.; Reed, G.; Biever, D.; et al. Genetically improved potatoes: Protection from damage by Colorado potato beetles. Plant Mol. Biol. 1993, 22, 313–321. [Google Scholar] [CrossRef]
- Whalon, M.E.; Wingerd, B.A. Bt: Mode of action and use. Arch. Insect Biochem. Physiol. Publ. Collab. Entomol. Soc. Am. 2003, 54, 200–211. [Google Scholar] [CrossRef]
- Sexson, D.L.; Wyman, J.A. Effect of crop rotation distance on populations of Colorado potato beetle (Coleoptera: Chrysomelidae): Development of areawide Colorado potato beetle pest management strategies. J. Econ. Entomol. 2005, 98, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Christou, P.; Capell, T.; Kohli, A.; Gatehouse, J.A.; Gatehouse, A.M. Recent developments and future prospects in insect pest control in transgenic crops. Trends Plant Sci. 2006, 11, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Fischhoff, D.A.; Fuchs, R.L.; Lavrik, P.B.; McPherson, S.A.; Perlak, F.J. Insect Resistant Tomato and Potato Plants. U.S. Patent No. 5,495,071, 27 February 1996. [Google Scholar]
- Thomas, P.E.; Kaniewski, W.K.; Lawson, E.C. Reduced field spread of potato leafroll virus in potatoes transformed with the potato leafroll virus coat protein gene. Plant Dis. 1997, 81, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- Adang, M.J.; Brody, M.S.; Cardineau, G.; Eagan, N.; Roush, R.T.; Shewmaker, C.K.; Jones, A.; Oakes, J.V.; McBride, K.E. The reconstruction and expression of a Bacillus thuringiensis cryIIIA gene in protoplasts and potato plants. Plant Mol. Biol. 1993, 21, 1131–1145. [Google Scholar] [CrossRef] [PubMed]
- Haffani, Y.Z.; Overney, S.; Yelle, S.; Bellemare, G.; Belzile, F.J. Premature polyadenylation contributes to the poor expression of the Bacillus thuringiensis cry3Ca1 gene in transgenic potato plants. Mol. Gen. Genet. 2000, 264, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Naimov, S.; Weemen-Hendriks, M.; Dukiandjiev, S.; de Maagd, R.A. Bacillus thuringiensis delta-endotoxin Cry1 hybrid proteins with increased activity against the Colorado potato beetle. Appl. Environ. Microbiol. 2001, 67, 5328–5330. [Google Scholar] [CrossRef] [Green Version]
- Meissle, M.; Romeis, J. Insecticidal activity of Cry3Bb1 expressed in Bt maize on larvae of the Colorado potato beetle, Leptinotarsa Decemlineata. Entomol. Exp. Appl. 2009, 131, 308–319. [Google Scholar] [CrossRef]
- Reed, G.L.; Jensen, A.S.; Riebe, J.; Head, G.; Duan, J.J. Transgenic Bt potato and conventional insecticides for Colorado potato beetle management: Comparative efficacy and non-target impacts. Entomol. Exp. Appl. 2001, 100, 89–100. [Google Scholar] [CrossRef]
- Keller, B.; Langenbruch, G.A. Control of coleopteran pests by Bacillus thuringiensis. In Bacillus Thuringiensis, an Environmental Biopesticide: Theory and Practice; John Wiley & Sons: New York, NY, USA, 1993; pp. 171–191. [Google Scholar]
- Kaniewski, W.K.; Thomas, P.E. The potato story. J. Agrobiotechnol. Manag. Econ. 2004, 7, 8. [Google Scholar]
- Alyokhin, A.V.; Ferro, D.N. Relative fitness of Colorado potato beetle (Coleoptera: Chrysomelidae) resistant and susceptible to the Bacillus thuringiensis Cry3A toxin. J. Econ. Entomol. 1999, 92, 510–515. [Google Scholar] [CrossRef] [Green Version]
- Hoy, C.W. Colorado potato beetle resistance management strategies for transgenic potatoes. Am. J. Potato Res. 1999, 76, 215–219. [Google Scholar] [CrossRef]
- Whalon, M.E.; Ferro, D.N. Bt-potato resistance management. In Now or Never: Serious New Plans to Save a Natural Pest Control; Union of Concerned Scientists: Cambridge, MA, USA, 1998. [Google Scholar]
- Thornton, M. The rise and fall of NewLeaf potatoes. NABC Rep. 2003, 15, 235–243. [Google Scholar]
- Zhao, J.Z.; Bishop, B.A.; Grafius, E.J. Inheritance and synergism of resistance to imidacloprid in the Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2000, 93, 1508–1514. [Google Scholar] [CrossRef] [PubMed]
- Olson, E.R.; Dively, G.P.; Nelson, J.O. Baseline susceptibility to imidacloprid and cross resistance patterns in Colorado potato beetle (Coleoptera: Chrysomelidae) populations. J. Econ. Entomol. 2000, 93, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Mota-Sanchez, D.; Hollingworth, R.M.; Grafius, E.J.; Moyer, D.D. Resistance and cross-resistance to neonicotinoid insecticides and spinosad in the Colorado potato beetle, Leptinotarsa decemlineata (Say)(Coleoptera: Chrysomelidae). Pest Manag. Sci. 2006, 62, 30–37. [Google Scholar] [CrossRef]
- Alyokhin, A.; Dively, G.; Patterson, M.; Castaldo, C.; Rogers, D.; Mahoney, M.; Wollam, J. Resistance and cross-resistance to imidacloprid and thiamethoxam in the Colorado potato beetle Leptinotarsa Decemlineata. Pest Manag. Sci. 2007, 63, 32–41. [Google Scholar] [CrossRef]
- Spooner, D.M.; Bamberg, J.B. Potato genetic resources: Sources of resistance and systematics. Am. Potato J. 1994, 71, 325–337. [Google Scholar] [CrossRef]
- Balbyshev, N.F.; Lorenzen, J.H. Hypersensitivity and egg drop: A novel mechanism of host plant resistance to Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1997, 90, 652–657. [Google Scholar] [CrossRef]
- Lorenzen, J.H.; Balbyshev, N.F.; Lafta, A.M.; Casper, H.; Tian, X.; Sagredo, B. Resistant potato selections contain leptine and inhibit development of the Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2001, 94, 1260–1267. [Google Scholar] [CrossRef]
- Coombs, J.J.; Douches, D.S.; Li, W.; Grafius, E.J.; Pett, W.L. Combining engineered (Bt-cry3A) and natural resistance mechanisms in potato for control of Colorado potato beetle. J. Am. Soc. Hortic. Sci. 2002, 127, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Mansoor, S.; Amin, I.; Hussain, M.; Zafar, Y.; Briddon, R.W. Engineering novel traits in plants through RNA interference. Trends Plant Sci. 2006, 11, 559–565. [Google Scholar] [CrossRef] [PubMed]
- He, W.W.; Xu, S.J.; Xu, L.T.; Zhang, J. RNA interference in Colorado potato beetle (Leptinotarsa decemlineata): A potential strategy for pest control. J. Integr. Agric. 2020, 19, 428–437. [Google Scholar]
- Zhang, J.; Khan, S.A.; Heckel, D.G.; Bock, R. Next-generation insect-resistant plants: RNAi-mediated crop protection. Trends Biotechnol. 2017, 35, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, H.C.; Miao, X.X. Feasibility, limitation and possible solutions of RNAi-based technology for insect pest control. Insect Sci. 2013, 20, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Dowling, D.P.; Miles, Z.D.; Köhrer, C.; Maiocco, S.J.; Elliott, S.J.; Bandarian, V.; Drennan, C.L. Molecular basis of cobalamin-dependent RNA modification. Nucleic Acids Res. 2016, 44, 9965–9976. [Google Scholar] [CrossRef] [Green Version]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Swevers, L.; Smagghe, G. Use of RNAi for control of insect crop pests. In Arthropod-Plant Interactions, 1st ed.; Smagghe, G., Diaz, I., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 177–197. [Google Scholar]
- Kumar, A.; Congiu, L.; Lindström, L.; Piiroinen, S.; Vidotto, M.; Grapputo, A. Sequencing, de novo assembly and annotation of the Colorado potato beetle, Leptinotarsa decemlineata, transcriptome. PLoS ONE 2014, 9, e86012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoville, S.D.; Chen, Y.H.; Andersson, M.N.; Benoit, J.B.; Bhandari, A.; Bowsher, J.H.; Brevik, K.; Cappelle, K.; Chen, M.J.M.; Childers, A.K.; et al. A model species for agricultural pest genomics: The genome of the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Sci. Rep. 2018, 8, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Xu, J.; Palli, R.; Ferguson, J.; Palli, S.R. Ingested RNA interference for managing the populations of the Colorado potato beetle, Leptinotarsa Decemlineata. Pest Manag. Sci. 2011, 67, 175–182. [Google Scholar] [CrossRef]
- Zhou, L.T.; Jia, S.; Wan, P.J.; Kong, Y.; Guo, W.C.; Ahmat, T.; Li, G.Q. RNA interference of a putative S-adenosyl-L-homocysteine hydrolase gene affects larval performance in Leptinotarsa decemlineata (Say). J. Insect Physiol. 2013, 59, 1049–1056. [Google Scholar] [CrossRef]
- Wan, Y.; Qu, K.; Zhang, Q.C.; Flynn, R.A.; Manor, O.; Ouyang, Z.; Zhang, J.; Spitale, R.C.; Snyder, M.P.; Segal, E.; et al. Landscape and variation of RNA secondary structure across the human transcriptome. Nature 2014, 505, 706–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaddelapati, S.C.; Kalsi, M.; Roy, A.; Palli, S.R. Cap’n’collar C regulates genes responsible for imidacloprid resistance in the Colorado potato beetle, Leptinotarsa decemlineata. Insect Biochem. Mol. Biol. 2018, 99, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Liu, X.P.; Wan, P.J.; Shi, X.Q.; Guo, W.C.; Li, G.Q. The P450 enzyme Shade mediates the hydroxylation of ecdysone to 20-hydroxyecdysone in the Colorado potato beetle, Leptinotarsa decemlineata. Insect Mol. Biol. 2014, 23, 632–643. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Campuzano, C.; Martínez-Ramírez, A.C.; Contreras, E.; Rausell, C.; Real, M.D. Prohibitin, an essential protein for Colorado potato beetle larval viability, is relevant to Bacillus thuringiensis Cry3Aa toxicity. Pestic. Biochem. Physiol. 2013, 107, 299–308. [Google Scholar] [CrossRef]
- Wan, P.J.; Fu, K.Y.; Lü, F.G.; Guo, W.C.; Li, G.Q. Knockdown of a putative alanine aminotransferase gene affects amino acid content and flight capacity in the Colorado potato beetle Leptinotarsa decemlineata. Amino Acids 2015, 47, 1445–1454. [Google Scholar] [CrossRef]
- Wan, P.J.; Fu, K.Y.; Lü, F.G.; Wang, X.X.; Guo, W.C.; Li, G.Q. Knocking down a putative Δ1-pyrroline-5-carboxylate dehydrogenase gene by RNA interference inhibits flight and causes adult lethality in the Colorado potato beetle Leptinotarsa decemlineata (Say). Pest Manag. Sci. 2015, 71, 1387–1396. [Google Scholar] [CrossRef]
- Hussain, T.; Aksoy, E.; Çalışkan, M.E.; Bakhsh, A. Transgenic potato lines expressing hairpin RNAi construct of molting-associated EcR gene exhibit enhanced resistance against Colorado potato beetle (Leptinotarsa decemlineata, Say). Transgenic Res. 2019, 28, 151–164. [Google Scholar] [CrossRef]
- Liu, X.P.; Fu, K.Y.; Lü, F.G.; Meng, Q.W.; Guo, W.C.; Li, G.Q. Involvement of FTZ-F1 in the regulation of pupation in Leptinotarsa decemlineata (Say). Insect Biochem. Mol. Biol. 2014, 55, 51–60. [Google Scholar] [CrossRef]
- Fu, K.Y.; Guo, W.C.; Ahmat, T.; Li, G.Q. Knockdown of a nutrient amino acid transporter gene LdNAT1 reduces free neutral amino acid contents and impairs Leptinotarsa decemlineata pupation. Sci. Rep. 2015, 5, 18124. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Khan, S.A.; Hasse, C.; Ruf, S.; Heckel, D.G.; Bock, R. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science 2015, 347, 991–994. [Google Scholar] [CrossRef]
- Lü, F.G.; Fu, K.Y.; Guo, W.C.; Li, G.Q. Characterization of two juvenile hormone epoxide hydrolases by RNA interference in the Colorado potato beetle. Gene 2015, 570, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.C.; Liu, X.P.; Fu, K.Y.; Shi, J.F.; Lü, F.G.; Li, G.Q. Functions of nuclear receptor HR3 during larval-pupal molting in Leptinotarsa decemlineata (Say) revealed by in vivo RNA interference. Insect Biochem. Mol. Biol. 2015, 63, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.F.; Fu, J.; Mu, L.L.; Guo, W.C.; Li, G.Q. Two Leptinotarsa uridine diphosphate N-acetylglucosamine pyrophosphorylases are specialized for chitin synthesis in larval epidermal cuticle and midgut peritrophic matrix. Insect Biochem. Mol. Biol. 2016, 68, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.F.; Mu, L.L.; Chen, X.; Guo, W.C.; Li, G.Q. RNA interference of chitin synthase genes inhibits chitin biosynthesis and affects larval performance in Leptinotarsa decemlineata (Say). Int. J. Biol. Sci. 2016, 12, 1319. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.F.; Xu, Q.Y.; Sun, Q.K.; Meng, Q.W.; Mu, L.L.; Guo, W.C.; Li, G.Q. Physiological roles of trehalose in Leptinotarsa larvae revealed by RNA interference of trehalose-6-phosphate synthase and trehalase genes. Insect Biochem. Mol. Biol. 2016, 77, 52–68. [Google Scholar] [CrossRef]
- Guo, W.C.; Liu, X.P.; Fu, K.Y.; Shi, J.F.; Lü, F.G.; Li, G.Q. Nuclear receptor ecdysone-induced protein 75 is required for larval–pupal metamorphosis in the Colorado potato beetle Leptinotarsa decemlineata (Say). Insect Mol. Biol. 2016, 25, 44–57. [Google Scholar] [CrossRef]
- Fu, K.Y.; Li, Q.; Zhou, L.T.; Meng, Q.W.; Lü, F.G.; Guo, W.C.; Li, G.Q. Knockdown of juvenile hormone acid methyl transferase severely affects the performance of Leptinotarsa decemlineata (Say) larvae and adults. Pest Manag. Sci. 2016, 72, 1231–1241. [Google Scholar] [CrossRef]
- Fu, K.Y.; Zhu, T.T.; Guo, W.C.; Ahmat, T.; Li, G.Q. Knockdown of a putative insulin-like peptide gene LdILP2 in Leptinotarsa decemlineata by RNA interference impairs pupation and adult emergence. Gene 2016, 581, 170–177. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, Y.; Zhang, S.; Deng, J.; Lu, M.; Zhang, L.; Zhang, J. Comparative analysis of the immune system of an invasive bark beetle, Dendroctonus valens, infected by an entomopathogenic fungus. Dev. Comp. Immunol. 2018, 88, 65–69. [Google Scholar] [CrossRef]
- Petek, M.; Coll, A.; Razinger, J.; Gruden, K. Validating the potential of double-stranded RNA targeting Colorado potato beetle mesh gene in laboratory and field trials. bioRxiv 2020. [Google Scholar] [CrossRef]
- Palli, S.R. RNA interference in Colorado potato beetle: Steps toward development of dsRNA as a commercial insecticide. Curr. Opin. Insect Sci. 2014, 6, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, S.; Taning, C.N.T.; Wei, D.; Smagghe, G. First report on CRISPR/Cas9-targeted mutagenesis in the Colorado potato beetle, Leptinotarsa Decemlineata. J. Insect Physiol. 2020, 121, 104013. [Google Scholar] [CrossRef]
- Whitten, M.M.; Facey, P.D.; Del Sol, R.; Fernández-Martínez, L.T.; Evans, M.C.; Mitchell, J.J.; Bodger, O.G.; Dyson, P.J. Symbiont-mediated RNA interference in insects. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taning, C.N.; Christiaens, O.; Li, X.; Swevers, L.; Casteels, H.; Maes, M.; Smagghe, G. Engineered flock house virus for targeted gene suppression through RNAi in fruit flies (Drosophila melanogaster) in vitro and in vivo. Front. Physiol. 2018, 9, 805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cagliari, D.; Avila dos Santos, E.; Dias, N.; Smagghe, G.; Zotti, M. Nontransformative strategies for RNAi in crop protection. In Modulating Gene Expression-Abridging the RNAi and CRISPR-Cas9 Technologies; IntechOpen: Rijeka, Croatia, 2019. [Google Scholar]
- Christiaens, O.; Dzhambazova, T.; Kostov, K.; Arpaia, S.; Joga, M.R.; Urru, I.; Sweet, J.; Smagghe, G. Literature review of baseline information on RNAi to support the environmental risk assessment of RNAi-based GM plants. EFSA Supporting Publ. 2018, 15, 1424E. [Google Scholar] [CrossRef]
- Burand, J.P.; Hunter, W.B. RNAi: Future in insect management. J. Invertebr. Pathol. 2013, 112, S68–S74. [Google Scholar] [CrossRef] [PubMed]
- Katoch, R.; Sethi, A.; Thakur, N.; Murdock, L.L. RNAi for insect control: Current perspective and future challenges. Appl. Biochem. Biotechnol. 2013, 171, 847–873. [Google Scholar] [CrossRef]
- Clark, J.M.; Lee, S.H.; Kim, H.J.; Yoon, K.S.; Zhang, A. DNA-based genotyping techniques for the detection of point mutations associated with insecticide resistance in Colorado potato beetle Leptinotarsa Decemlineata. Pest Manag. Sci. 2001, 57, 968–974. [Google Scholar] [CrossRef]
- Udalov, M.B.; Benkovskaya, G.V. Population genetics of the Colorado potato beetle: From genotype to phenotype. Russ. J. Genet. Appl. Res. 2011, 1, 321. [Google Scholar] [CrossRef]
- Grapputo, A.; Boman, S.; Lindstroem, L.; Lyytinen, A.; Mappes, J. The voyage of an invasive species across continents: Genetic diversity of North American and European Colorado potato beetle populations. Mol. Ecol. 2005, 14, 4207–4219. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The population biology of invasive species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Xing, C.; Schumacher, F.R.; Xing, G.; Lu, Q.; Wang, T.; Elston, R.C. December. Comparison of microsatellites, single-nucleotide polymorphisms (SNPs) and composite markers derived from SNPs in linkage analysis. BMC Genet. 2005, 6, S29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genissel, A.; Pastinen, T.; Dowell, A.; Mackay, T.F.; Long, A.D. No evidence for an association between common nonsynonymous polymorphisms in Delta and bristle number variation in natural and laboratory populations of Drosophila melanogaster. Genetics 2004, 166, 291–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coates, B.S.; Sumerford, D.V.; Miller, N.J.; Kim, K.S.; Sappington, T.W.; Siegfried, B.D.; Lewis, L.C. Comparative performance of single nucleotide polymorphism and microsatellite markers for population genetic analysis. J. Hered. 2009, 100, 556–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotsakiozi, P.; Evans, B.R.; Gloria-Soria, A.; Kamgang, B.; Mayanja, M.; Lutwama, J.; Le Goff, G.; Ayala, D.; Paupy, C.; Badolo, A.; et al. Population structure of a vector of human diseases: Aedes aegypti in its ancestral range, Africa. Ecol. Evol. 2018, 8, 7835–7848. [Google Scholar] [CrossRef] [Green Version]
- Saarman, N.P.; Opiro, R.; Hyseni, C.; Echodu, R.; Opiyo, E.A.; Dion, K.; Johnson, T.; Aksoy, S.; Caccone, A. The population genomics of multiple tsetse fly (Glossina fuscipes fuscipes) admixture zones in Uganda. Mol. Ecol. 2019, 28, 66–85. [Google Scholar] [CrossRef] [Green Version]
- Francischini, F.J.; Cordeiro, E.M.; de Campos, J.B.; Alves-Pereira, A.; Viana, J.P.G.; Wu, X.; Wei, W.; Brown, P.; Joyce, A.; Murua, G.; et al. Diatraea saccharalis history of colonization in the Americas. The case for human-mediated dispersal. PLoS ONE 2019, 14, e0220031. [Google Scholar] [CrossRef]
- Yadav, S.; Stow, A.J.; Dudaniec, R.Y. Detection of environmental and morphological adaptation despite high landscape genetic connectivity in a pest grasshopper (Phaulacridium vittatum). Mol. Ecol. 2019, 8, 3395–3412. [Google Scholar] [CrossRef]
- Brumfield, R.T.; Beerli, P.; Nickerson, D.A.; Edwards, S.V. The utility of single nucleotide polymorphisms in inferences of population history. Trends Ecol. Evol. 2003, 18, 249–256. [Google Scholar] [CrossRef]
- Morin, P.A.; Luikart, G.; Wayne, R.K. SNPs in ecology, evolution and conservation. Trends Ecol. Evol. 2004, 19, 208–216. [Google Scholar] [CrossRef]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity arrays: A solid state technology for sequence information independent genotyping. Nucleic Acids Res. 2001, 29, e25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nantoume, A.D.; Andersen, S.B.; Jensen, B.D. Genetic differentiation of watermelon landrace types in Mali revealed by microsatellite (SSR) markers. Genet. Resour. Crop Evol. 2013, 60, 2129–2141. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Annotation | Reference |
|---|---|---|
| VATPase, A, B, E | Vacuolar ATP synthase subunit | [61,65] |
| Sec23 | Protein transport protein sec23 | [65] |
| COPβ | Coatomer β-subunit | [65] |
| Actin | β-Actin | [65] |
| Prohibitin | Prohibitin protein | [70] |
| SAHase | S-adenosyl-L-homocysteine hydrolase | [66] |
| FTZ-F1 | Nuclear receptor type transcription factor that responses to 20-hydroxyecdysone | [74] |
| shd | Ecdysone 20-monooxygenase | [69] |
| NAT1 | Nutrient amino acid transporter | [75] |
| Actin | β-Actin | [76] |
| JHEH | Juvenile hormone epoxide hydrolase | [77] |
| alt | Alanine aminotransferase | [71] |
| p5cdh | Δ1-pyrroline-5-carboxylate dehydrogenase | [72] |
| HR3 | Nuclear receptor that early-late responses to 20-Hydroxyecdysone | [78] |
| UAP | Uridine diphosphate N-acetylglucosamine pyrophosphorylase | [79] |
| ChS | Chitin synthase | [80] |
| TPS and TREs | Trehalose biosynthesis and degradation | [81] |
| E75 | Ecdysone-induced protein 75 | [82] |
| JHAMT | Juvenile hormone acid methyltransferase | [83] |
| ILP2 | Putative insulin-like peptide | [84] |
| HR4 | ecdysteroidogenesis and mediates 20-hydroxyecdysone signaling during larval-pupal metamorphosis | [85] |
| CncC | Xenobiotic transcription factor | [69] |
| EcR | Ecdysone receptor | [73] |
| Mesh | gut-membrane-associated protein | [86] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadoić Balaško, M.; Mikac, K.M.; Bažok, R.; Lemic, D. Modern Techniques in Colorado Potato Beetle (Leptinotarsa decemlineata Say) Control and Resistance Management: History Review and Future Perspectives. Insects 2020, 11, 581. https://doi.org/10.3390/insects11090581
Kadoić Balaško M, Mikac KM, Bažok R, Lemic D. Modern Techniques in Colorado Potato Beetle (Leptinotarsa decemlineata Say) Control and Resistance Management: History Review and Future Perspectives. Insects. 2020; 11(9):581. https://doi.org/10.3390/insects11090581
Chicago/Turabian StyleKadoić Balaško, Martina, Katarina M. Mikac, Renata Bažok, and Darija Lemic. 2020. "Modern Techniques in Colorado Potato Beetle (Leptinotarsa decemlineata Say) Control and Resistance Management: History Review and Future Perspectives" Insects 11, no. 9: 581. https://doi.org/10.3390/insects11090581