
Assessing the Ecophysiology of Methanogens in the Context of Recent Astrobiological and Planetological Studies

 , ,
, ,
Abstract
:1. Introduction
- (1)
- from hydrogen (H) and carbon dioxide (CO) or from carbon monoxide (CO) or from formate (HC)
- (2)
- from methanol, methylamines, methanethiol or methylsulfide
- (3)
- from acetate
- (4)
- or from methanol (CHOH), methylamines and methylsulfide as methyl group donor and H as electron source.

2. Ecophysiological Characteristics of Methanogens
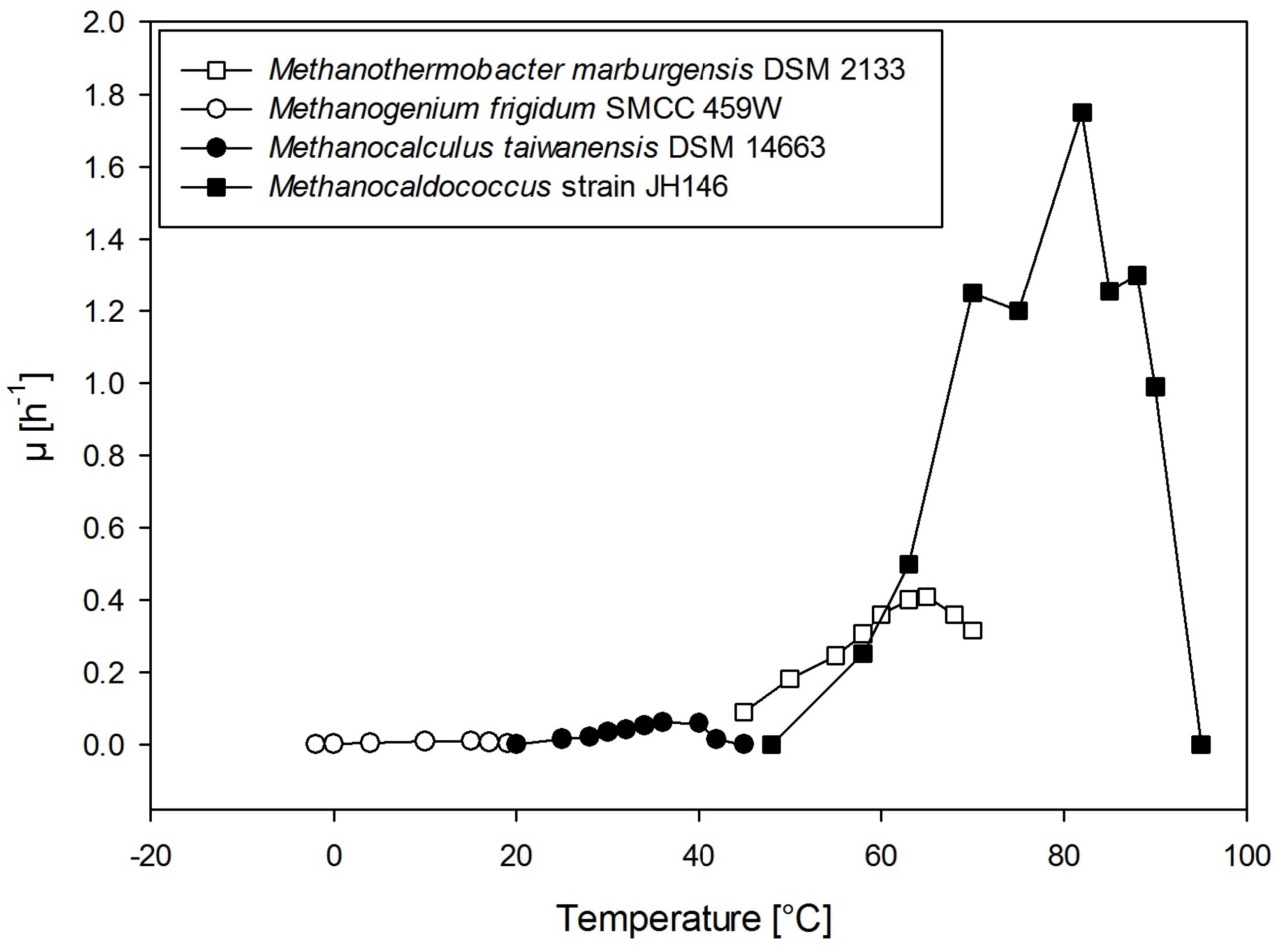
2.1. Temperature


| Strain | Temp. [C] | pH | Ref. | ||||
|---|---|---|---|---|---|---|---|
| min. | opt | max | min. | opt. | max. | ||
| Methanospirillum psychrodurum | 4 | 25 | 32 | 6.5 | 7 | 8 | [92] |
| Methanosarcina baltica | 3 | 21 | 28 | 6.3 | 7.2 | 7.5 | [32] |
| Methanosarcina lacustris | 1 | 25 | 35 | 4.5 | 7 | 8.5 | [93] |
| Methanolobus psychrophilus | 0 | 18 | 25 | 6 | 7–7.2 | 8 | [94] |
| Methanogenium marinum | 5 | 25 | 25 | 5.5 | 6–6.6 | 7.7 | [95] |
| Methanogenium frigidum | 0 | 15 | 17 | 6.3 | 7.5–7.9 | 8 | [33] |
| Methanohalobium evestigatum | 50 | n.a. | n.a. | n.a. | 7.4 | n.a. | [96] |
| Methanogenium cariaci | 15 | 20–25 | 35 | 6 | 6.8–7.2 | 7.5 | [97] |
| Methanogenium boonei | 5 | 19.4 | 25.6 | 6.4 | n.a. | 7.8 | [98] |
| Methanoculleus marisnigri | 15 | 20–25 | 48 | 6 | 6.2–6.6 | 7.6 | [99] |
| Methanoculleus chikugoensis | 15 | 25 | 40 | 6.7 | 6.7–7.2 | 8 | [100] |
| Methanococcoides alaskense | 2.3 | 23.6 | 28.4 | 6.3 | n.a. | 7.5 | [101] |
| Methanococcoides burtonii | 1.7 | 23.4 | 29.5 | 6.8 | n.a. | 8.2 | [102] |
| Methanospirillum stamsii | 5 | 20–30 | 37 | 6.0 | 7.0–7.5 | 10 | [31] |
| Methanosarcina soligelidi | 0 | 28 | 54 | 4.8 | 7.8 | 9.9 | [34] |
| Candidatus “Methanoflorens stordalenmirensis” | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | [36] |
2.2. Pressure
2.3. pH
| Strain | Temp. [C] | pH | Ref. | ||||
|---|---|---|---|---|---|---|---|
| min. | opt. | max | min. | opt. | max. | ||
| Methanospirillum stamsii | 5 | 20–30 | 37 | 6 | 7–7.5 | 10 | [31] |
| Methanocalculus natronophilus | 14 | 30–37 | 45 | 8 | 9–9.5 | 10.2 | [121] |
| Methanospirillum hungatei | 20 | 37–45 | 50 | 6.5 | 7–9 | 10 | [12] |
| Methanobrevibacter millerae | 33 | 36–42 | 43 | 5.5 | 7–8 | 10 | [122] |
| Methanobrevibacter olleyae | 28 | 28–42 | 42 | 6 | 7.5 | 10 | [122] |
| Methanotorris igneus | 45 | 88 | 91 | 5 | 5.7 | 7.5 | [123] |
| Methanosphaerula palustris | 14 | 30 | 35 | 4.8 | 5.5 | 6.4 | [120] |
| Methanoregula boonei | 10 | 35–37 | 40 | 4.5 | 5.1 | 5.5 | [98] |
2.4. Osmolarity
2.5. Ionizing and UV Radiation
2.6. Desiccation
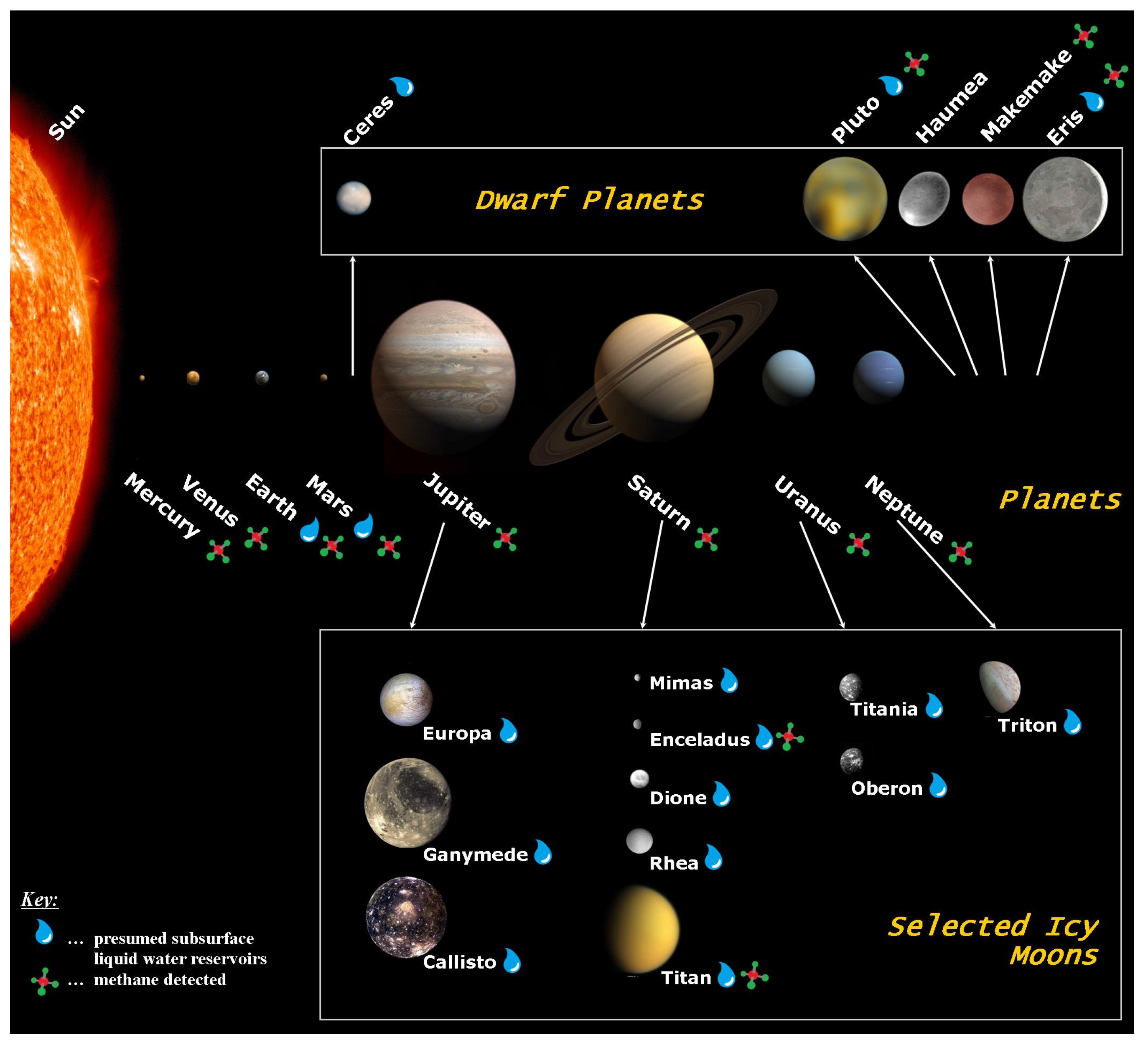
3. Known and Potential Habitats for Methanogens in the Solar System

| Host Planet | R [km] | ρ [kg m] | T [K] | Atmosphere | Water/Core | |||
|---|---|---|---|---|---|---|---|---|
| Moon | Earth | 1737.4 ± 1 [150] | 3344 ± 5 [151] | 26 [152] | 220 [153] | 390 | no | no |
| Mars | - | 3389.50 ± 0.2 [150] | 5.5134 ± 0.0006 [154] | 150 [155] | 215 [155] | 290 [155] | yes [155] | - |
| Ceres | - | 476.2± 1.7 [150] | 2077 ± 36 [156] | n.a. | n.a. | n.a. | no | no (?) |
| Ganymede | Jupiter | 2631.2 ± 1.7 [150] | 1942 ± 5 [151] | 70 [157] | 109 [153] | 152 [157] | no | no/yes |
| Callisto | Jupiter | 2410.3 ± 1.5 [150] | 1834 ± 4 [151] | 80±5 [158] | 134 [153] | 165 [158] | yes (thin) | no |
| Europa | Jupiter | 1560.8 ± 0.3 [150] | 3013 ± 5 [151] | 50 [153] | 103 [153] | n.a. | yes (thin) | yes |
| Titan | Saturn | 2574.73 ± 0.09 [150] | 1882 ± 1 [151] | n.a. | 94 [159] | n.a. | yes | no (?) |
| Enceladus | Saturn | 252.1 ± 0.2 [160] | 1609 ± 5 [160] | 32.9 [161] | 75 [161] | ≥157 [162] | local [162] | yes (?) [163] |
| Dione | Saturn | 561.4 ± 0.4 [150] | 1476 ± 4 [151] | n.a. | n.a. | n.a. | no | n.a. |
| Mimas | Saturn | 198.2 ± 0.4 [150] | 1150 ± 4 [151] | n.a. | n.a. | n.a. | no | no |
| Triton | Neptune | 1352.6 ± 2.4 [150] | 2059 ± 5 [151] | n.a. | 39 [155] | n.a. | yes (thin) | yes (?) |
| Pluto | - | 1195 ± 5 [150] | 2030±60 [164] | 35 [165] | 44 | 55 [165] | yes (thin) | yes (?) |
3.1. Earth
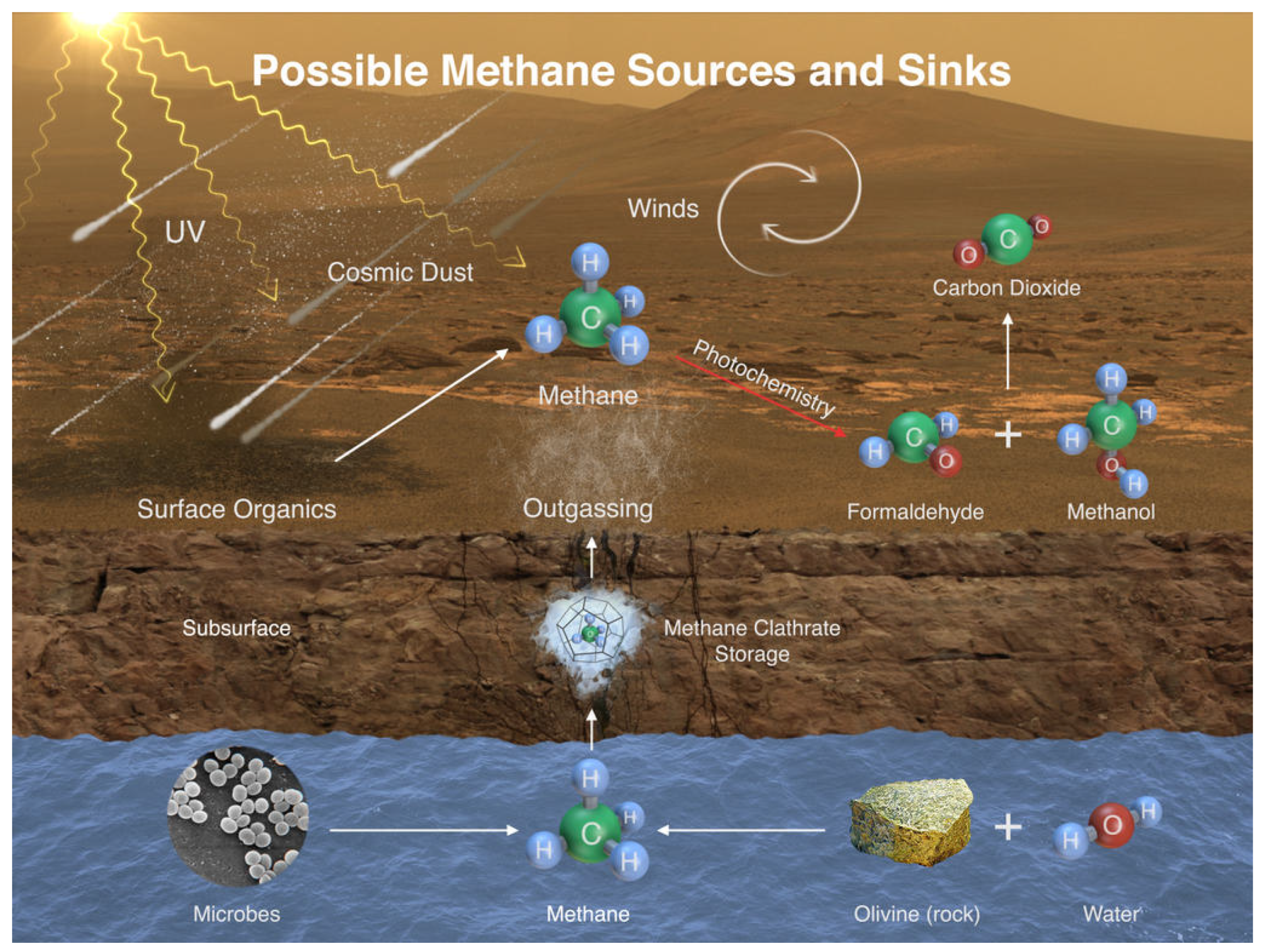
3.2. Mars

3.3. Potential Habitable Moons and Small Icy Bodies
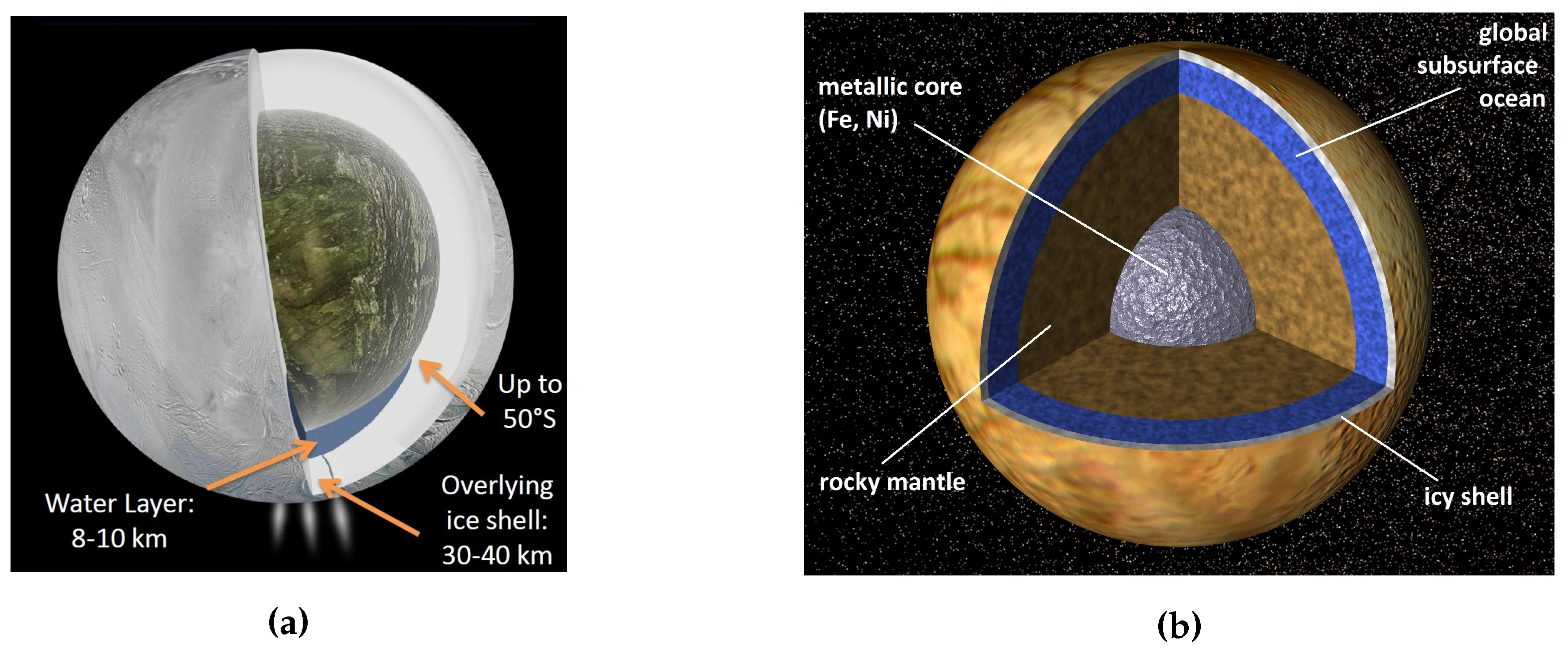
3.3.1. Enceladus

3.3.2. Europa
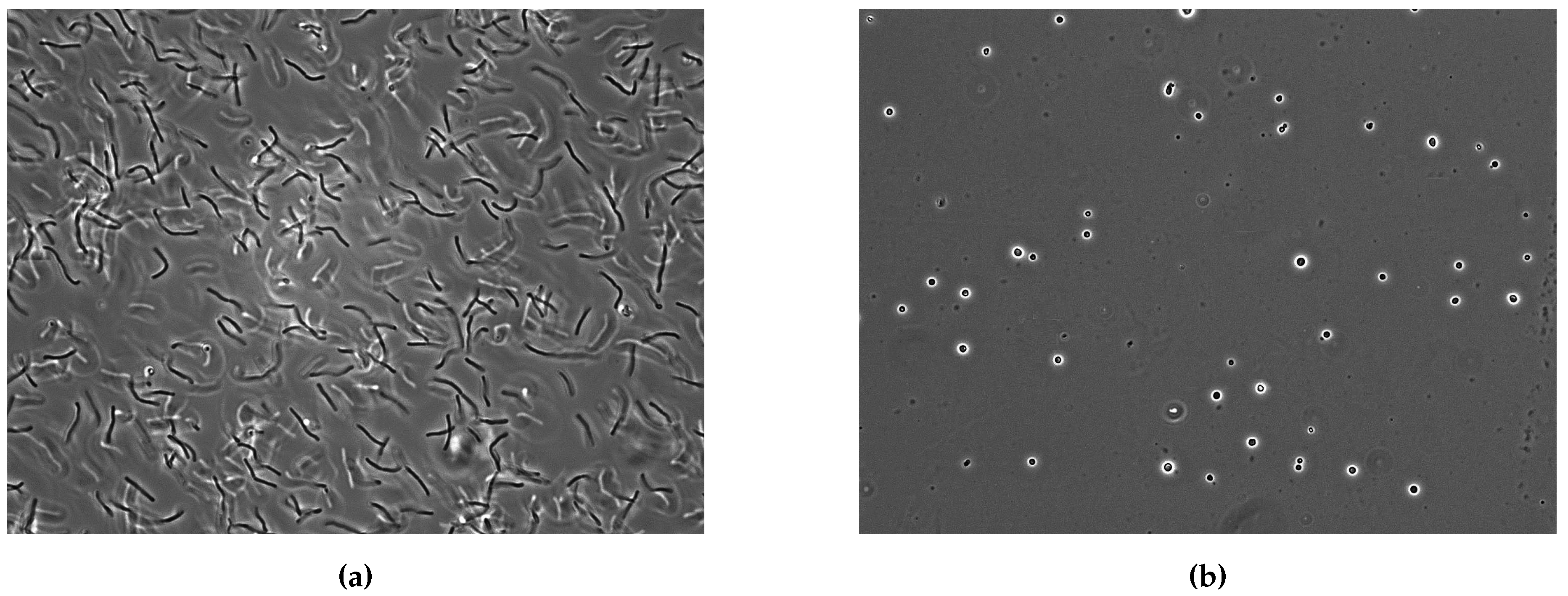
3.3.3. Titan

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3.3.4. Other Icy Moons
Ganymede
Mimas and Dione
Callisto
Triton
Rhea, Oberon, Titania
3.3.5. Dwarf Planets and Small Solar System Bodies
4. Simulating Extraterrestrial Conditions
4.1. Mars-Like Conditions
4.2. Icy Moon-Like Conditions
4.3. Terrestrial Analogues for Solar System Objects
5. Conclusions
5.1. Discussion
5.2. Outlook
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Niemann, H.B.; Atreya, S.K.; Bauer, S.J.; Carignan, G.R.; Demick, J.E.; Frost, R.L.; Gautier, D.; Haberman, J.A.; Harpold, D.N.; Hunten, D.M.; et al. The abundances of constituents of Titan’s atmosphere from the GCMS instrument on the Huygens probe. Nature 2005, 438, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Waite, J.H.; Brockwell, T.; Lewis, W.S.; Magee, B.; McKinnon, W.B.; Mousis, O.; Bouquet, A. Enceladus Plume Composition. LPI Contrib. 2014, 1774, 4013. [Google Scholar]
- Formisano, V.; Atreya, S.; Encrenaz, T.; Ignatiev, N.; Giuranna, M. Detection of Methane in the Atmosphere of Mars. Science 2004, 306, 1758–1761. [Google Scholar] [CrossRef] [PubMed]
- Mumma, M.J.; Villanueva, G.L.; Novak, R.E.; Hewagama, T.; Bonev, B.P.; DiSanti, M.A.; Mandell, A.M.; Smith, M.D. Strong Release of Methane on Mars in Northern Summer 2003. Science 2009, 323, 1041–1045. [Google Scholar] [CrossRef] [PubMed]
- Webster, C.R.; Mahaffy, P.R.; Atreya, S.K.; Flesch, G.J.; Mischna, M.A.; Meslin, P.Y.; Farley, K.A.; Conrad, P.G.; Christensen, L.E.; Pavlov, A.A.; et al. Mars methane detection and variability at Gale crater. Science 2015, 347, 415–417. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Whitman, W.B. Metabolic, Phylogenetic, and Ecological Diversity of the Methanogenic Archaea. Ann. N. Y. Acad. Sci. 2008, 1125, 171–189. [Google Scholar] [CrossRef] [PubMed]
- There are also (aerobic) marine microorganisms known to produce CH4 from methylphosphonic acid [266,267,268].
- Offre, P.; Spang, A.; Schleper, C. Archaea in Biogeochemical Cycles. Annu. Rev. Microbiol. 2013, 67, 437–457. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.N.; Parks, D.H.; Chadwick, G.L.; Robbins, S.J.; Orphan, V.J.; Golding, S.D.; Tyson, G.W. Methane metabolism in the archaeal phylum Bathyarchaeota revealed by genome-centric metagenomics. Science 2015, 350, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Thauer, R.K.; Kaster, A.K.; Seedorf, H.; Buckel, W.; Hedderich, R. Methanogenic archaea: Ecologically relevant differences in energy conservation. Nat. Rev. Microbiol. 2008, 6, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Borrel, G.; O’Toole, P.W.; Harris, H.M.B.; Peyret, P.; Brugère, J.F.; Gribaldo, S. Phylogenomic Data Support a Seventh Order of Methylotrophic Methanogens and Provide Insights into the Evolution of Methanogenesis. Genome Biol. Evol. 2013, 5, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Iino, T.; Tamaki, H.; Tamazawa, S.; Ueno, Y.; Ohkuma, M.; Suzuki, K.i.; Igarashi, Y.; Haruta, S. Candidatus Methanogranum caenicola: A Novel Methanogen from the Anaerobic Digested Sludge, and Proposal of Methanomassiliicoccaceae fam. nov. and Methanomassiliicoccales ord. nov., for a Methanogenic Lineage of the Class Thermoplasmata. Microbes Environ. 2013, 28, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Rittmann, S.; Seifert, A.; Herwig, C. Essential prerequisites for successful bioprocess development of biological CH4 production from CO2 and H2. Crit. Rev. Biotechnol. 2015, 35, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Ferry, J.G. How to make a living by exhaling methane. Annu. Rev. Microbiol. 2010, 64, 453–473. [Google Scholar] [CrossRef] [PubMed]
- Oelgeschläger, E.; Rother, M. Influence of carbon monoxide on metabolite formation in Methanosarcina acetivorans. FEMS Microbiol. Lett. 2009, 292, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Rother, M.; Metcalf, W.W. Anaerobic growth of Methanosarcina acetivorans C2A on carbon monoxide: An unusual way of life for a methanogenic archaeon. Proc. Natl. Acad. Sci. USA 2004, 101, 16929–16934. [Google Scholar] [CrossRef] [PubMed]
- Brocks, J.J.; Logan, G.A.; Buick, R.; Summons, R.E. Archean Molecular Fossils and the Early Rise of Eukaryotes. Science 1999, 285, 1033–1036. [Google Scholar] [CrossRef] [PubMed]
- Ueno, Y.; Yamada, K.; Yoshida, N.; Maruyama, S.; Isozak, Y. Evidence from fluid inclusions for microbial methanogenesis in the early Archaean era. Nature 2006, 440, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Brochier-Armanet, C.; Forterre, P.; Gribaldo, S. Phylogeny and evolution of the Archaea: One hundred genomes later. Curr. Opin. Microbiol. 2011, 14, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Blank, C.E. Phylogenomic dating—The relative antiquity of archaeal metabolic and physiological traits. Astrobiology 2009, 9, 193–219. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Takahashi, A.; Mori, C.; Tamaki, H.; Mochimaru, H.; Nakamura, K.; Takamizawa, K.; Kamagata, Y. Methanothermobacter tenebrarum sp. nov., a hydrogenotrophic, thermophilic methanogen isolated from gas-associated formation water of a natural gas field. Int. J. Syst. Evol. Microbiol. 2013, 63, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Liu, X.; Dong, X. Methanosaeta harundinacea sp. nov., a novel acetate-scavenging methanogen isolated from a UASB reactor. Int. J. Syst. Evol. Microbiol. 2006, 56, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Lü, Z.; Lu, Y. Methanocella conradii sp. nov., a Thermophilic, Obligate Hydrogenotrophic Methanogen, Isolated from Chinese Rice Field Soil. PLoS ONE 2012, 7, e35279. [Google Scholar] [CrossRef] [PubMed]
- L’Haridon, S.; Reysenbach, A.L.; Banta, A.; Messner, P.; Schumann, P.; Stackebrandt, E.; Jeanthon, C. Methanocaldococcus indicus sp. nov., a novel hyperthermophilic methanogen isolated from the Central Indian Ridge. Int. J. Syst. Evol. Microbiol. 2003, 53, 1931–1935. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.J.; Leigh, J.A.; Mayer, F.; Woese, C.R.; Wolfe, R.S. Methanococcus jannaschii sp. nov., an extremely thermophilic methanogen from a submarine hydrothermal vent. Arch. Microbiol. 1983, 136, 254–261. [Google Scholar] [CrossRef]
- Jiang, B.; Parshina, S.N.; Doesburg, W.V.; Lomans, B.P.; Stams, A.J.M. Methanomethylovorans thermophila sp. nov., a thermophilic, methylotrophic methanogen from an anaerobic reactor fed with methanol. Int. J. Syst. Evol. Microbiol. 2005, 55, 2465–2470. [Google Scholar] [CrossRef] [PubMed]
- Jeanthon, C.; L’Haridon, S.; Reysenbach, A.L.; Vernet, M.; Messner, P.; Sleytr, U.B.; Prieur, D. Methanococcus infernus sp. nov., a novel hyperthermophilic lithotrophic methanogen isolated from a deep-sea hydrothermal vent. Int. J. Syst. Bacteriol. 1998, 48 Pt 3, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Jeanthon, C.; L’Haridon, S.; Reysenbach, A.L.; Corre, E.; Vernet, M.; Messner, P.; Sleytr, U.B.; Prieur, D. Methanococcus vulcanius sp. nov., a novel hyperthermophilic methanogen isolated from East Pacific Rise, and identification of Methanococcus sp. DSM 4213T as Methanococcus fervens sp. nov. Int. J. Syst. Bacteriol. 1999, 49, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Qiu, T.L.; Yin, X.B.; Wu, X.L.; Hu, G.Q.; Deng, Y.; Zhang, H. Methermicoccus shengliensis gen. nov., sp. nov., a thermophilic, methylotrophic methanogen isolated from oil-production water, and proposal of Methermicoccaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 2964–2969. [Google Scholar] [CrossRef] [PubMed]
- Parshina, S.N.; Ermakova, A.V.; Bomberg, M.; Detkova, E.N. Methanospirillum stamsii sp. nov., a psychrotolerant, hydrogenotrophic, methanogenic archaeon isolated from an anaerobic expanded granular sludge bed bioreactor operated at low temperature. Int. J. Syst. Evol. Microbiol. 2014, 64, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Von Klein, D.v.; Arab, H.; Völker, H.; Thomm, M. Methanosarcina baltica, sp. nov., a novel methanogen isolated from the Gotland Deep of the Baltic Sea. Extremophiles 2002, 6, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Franzmann, P.D.; Liu, Y.; Balkwill, D.L.; Aldrich, H.C.; Macario, E.C.D.; Boone, D.R. Methanogenium frigidum sp. nov., a Psychrophilic, H2-Using Methanogen from Ace Lake, Antarctica. Int. J. Syst. Bacteriol. 1997, 47, 1068–1072. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.; Schirmack, J.; Ganzert, L.; Morozova, D.; Mangelsdorf, K. Methanosarcina soligelidi sp. nov., a desiccation- and freeze-thaw-resistant methanogenic archaeon from a Siberian permafrost-affected soil. Int. J. Syst. Evol. Microbiol. 2013, 63, 2986–2991. [Google Scholar] [CrossRef] [PubMed]
- Schirmack, J.; Mangelsdorf, K.; Ganzert, L.; Sand, W.; Hillebrand-Voiculescu, A.; Wagner, D. Methanobacterium movilense sp. nov., a hydrogenotrophic, secondary-alcohol-utilizing methanogen from the anoxic sediment of a subsurface lake. Int. J. Syst. Evol. Microbiol. 2014, 64, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Mondav, R.; Woodcroft, B.J.; Kim, E.H.; McCalley, C.K.; Hodgkins, S.B.; Crill, P.M.; Chanton, J.; Hurst, G.B.; VerBerkmoes, N.C.; Saleska, S.R.; et al. Discovery of a novel methanogen prevalent in thawing permafrost. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittmann, S.; Holubar, P. Rapid extraction of total RNA from an anaerobic sludge biocoenosis. Folia Microbiol. 2014, 59, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R. Cold-adapted archaea. Nat. Rev. Microbiol. 2006, 4, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.; Kurr, M.; Jannasch, H.W.; Stetter, K.O. A novel group of abyssal methanogenic archaebacteria (Methanopyrus) growing at 110 ∘C. Nature 1989, 342, 833–834. [Google Scholar] [CrossRef]
- Rittmann, S.K.-M.R.; Seifert, A.; Herwig, C. Quantitative analysis of media dilution rate effects on Methanothermobacter marburgensis grown in continuous culture on H2 and CO2. Biomass Bioenergy 2012, 36, 293–301. [Google Scholar] [CrossRef]
- Rittmann, S. K.-M. R. A critical assessment of microbiological biogas to biomethane upgrading systems. In Biogas Science and Technology; Guebitz, G.M., Ed.; Advances in Biochemical Engineering/Biotechnology; Springer International Publishing: New York, NY, USA, 2015. [Google Scholar]
- Seifert, A.H.; Rittmann, S.; Bernacchi, S.; Herwig, C. Method for assessing the impact of emission gasses on physiology and productivity in biological methanogenesis. Bioresour. Technol. 2013, 136, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Seifert, A.H.; Rittmann, S.; Herwig, C. Analysis of process related factors to increase volumetric productivity and quality of biomethane with Methanothermobacter marburgensis. Appl. Energy 2014, 132, 155–162. [Google Scholar] [CrossRef]
- Bernacchi, S.; Rittmann, S.; Seifert, A.H.; Krajete, A.; Herwig, C. Experimental methods for screening parameters influencing the growth to product yield (Y(x/CH4)) of a biological methane production (BMP) process performed with Methanothermobacter marburgensis. AIMS Bioeng. 2014, 1, 72–86. [Google Scholar] [CrossRef]
- Albers, S.V.; Meyer, B.H. The archaeal cell envelope. Nat. Rev. Microbiol. 2011, 9, 414–426. [Google Scholar] [CrossRef] [PubMed]
- UWr Wydział Biotechnologii. Methanogens Database. 2013. Available online: http://metanogen.biotech.uni.wroc.pl/ (accessed on 13 November 2015).
- Leibniz-Institut DSMZ. Catalogue of Microorganisms. 2015. Available online: http://www.dsmz.de/catalogues/catalogue-microorganisms.html (accessed on 13 November 2015).
- Schönheit, P.; Moll, J.; Thauer, R.K. Nickel, cobalt, and molybdenum requirement for growth of Methanobacterium thermoautotrophicum. Arch. Microbiol. 1979, 123, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Bonacker, L.G.; Baudner, S.; Mörschel, E.; Böcher, R.; Thauer, R.K. Properties of the two isoenzymes of methyl-coenzyme M reductase in Methanobacterium thermoautotrophicum. Eur. J. Biochem./FEBS 1993, 217, 587–595. [Google Scholar] [CrossRef]
- Wang, M.; Tomb, J.F.; Ferry, J.G. Electron transport in acetate-grown Methanosarcina acetivorans. BMC Microbiol. 2011, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Kaster, A.K.; Moll, J.; Parey, K.; Thauer, R.K. Coupling of ferredoxin and heterodisulfide reduction via electron bifurcation in hydrogenotrophic methanogenic archaea. Proc. Natl. Acad. Sci. USA 2011, 108, 2981–2986. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S. Occurrence of Life Outside the Solar System. Am. Sci. 1959, 47, 397–402. [Google Scholar]
- Kasting, J.F.; Whitmire, D.P.; Reynolds, R.T. Habitable Zones around Main Sequence Stars. Icarus 1993, 101, 108–128. [Google Scholar] [CrossRef] [PubMed]
- Leitner, J.J.; Schwarz, R.; Firneis, M.G.; Hitzenberger, R.; Neubauer, D. Generalizing Habitable Zones in Exoplanetary Systems—The Concept of the Life Supporting Zone. LPI Contrib. 2010, 1538, 5255. [Google Scholar]
- The water snow line describes the critical distance from a protostar in a protoplanetary disk where it is cold enough that water condenses into solid ice grains. In the Solar System, this line lies approx. at a distance of 2.7 AU from the Sun [269].
- The Martian history is roughly divided into three main periods, namely Noachian, Hersperian, and the present period named Amazonian.
- Martínez, G.M.; Renno, N.O. Water and Brines on Mars: Current Evidence and Implications for MSL. Space Sci. Rev. 2013, 175, 29–51. [Google Scholar] [CrossRef]
- Kral, T.A.; Altheide, T.S.; Lueders, A.E.; Schuerger, A.C. Low pressure and desiccation effects on methanogens: Implications for life on Mars. Planet. Space Sci. 2011, 59, 264–270. [Google Scholar] [CrossRef]
- Kral, T.A.; Altheide, S.T. Methanogen survival following exposure to desiccation, low pressure and martian regolith analogs. Planet. Space Sci. 2013, 89, 167–171. [Google Scholar] [CrossRef]
- Mickol, R.L.; Kral, T.A. Approaching Martian Conditions: Methanogen Survival at Low Pressure. In Lunar and Planetary Institute Technical Report, Proceedings of the 45th Lunar and Planetary Science Conference, Woodlands, TX, USA, 17–21 March 2014; Volume 45, p. 1602.
- Kral, T.; Bekkum, C.; McKay, C. Growth of Methanogens on a Mars Soil Simulant. Origin. Life Evol. Biosph. 2004, 34, 615–626. [Google Scholar] [CrossRef]
- Mickol, R.L.; Waddell, W.H.; Kral, T.A. Methanogens as Models for Life on Mars. LPI Contrib. 2014, 1791, 1005. [Google Scholar]
- Peters, G.H.; Abbey, W.; Bearman, G.H.; Mungas, G.S.; Smith, J.A.; Anderson, R.C.; Douglas, S.; Beegle, L.W. Mojave Mars simulant-Characterization of a new geologic Mars analog. Icarus 2008, 197, 470–479. [Google Scholar] [CrossRef]
- Kral, T.A.; Altheide, T.S.; Lueders, A.E.; Goodhart, T.H.; Virden, B.T.; Birch, W.; Howe, K.L.; Gavin, P. Methanogens: A Model for Life on Mars. LPI Contrib. 2010, 1538, 5084. [Google Scholar]
- Goodhart, T.; Kral, T.A. The Effects of Perchlorate on Methane Production of Methanogens. LPI Contrib. 2010, 1538, 5524. [Google Scholar]
- Chastain, B.K.; Kral, T.A. Approaching Mars-like Geochemical Conditions in the Laboratory: Omission of Artificial Buffers and Reductants in a Study of Biogenic Methane Production on a Smectite Clay. Astrobiology 2010, 10, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Chastain, B.K.; Kral, T.A. Zero-valent iron on Mars: An alternative energy source for methanogens. Icarus 2010, 208, 198–201. [Google Scholar] [CrossRef]
- Morozova, D.; Möhlmann, D.; Wagner, D. Survival of Methanogenic Archaea from Siberian Permafrost under Simulated Martian Thermal Conditions. Orig. Life Evol. Biosph. 2007, 37, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirmack, J.; Böhm, M.; Brauer, C.; Löhmannsröben, H.G.; de Vera, J.P.; Möhlmann, D.; Wagner, D. Laser spectroscopic real time measurements of methanogenic activity under simulated Martian subsurface analog conditions. Planet. Space Sci. 2014, 98, 198–204. [Google Scholar] [CrossRef]
- Serrano, P.; Wagner, D.; Böttger, U.; de Vera, J.P.; Lasch, P.; Hermelink, A. Single-cell analysis of the methanogenic archaeon Methanosarcina soligelidi from Siberian permafrost by means of confocal Raman microspectrocopy for astrobiological research. Planet. Space Sci. 2014, 98, 191–197. [Google Scholar] [CrossRef]
- De Vera, J.P.P.; Böttger, U.; Fritz, J.; Weber, I.; Malaszkiewicz, J.; Serrano, P.; Meessen, J.; Ott, S.; Wagner, D.; Hübers, H.W. Detection of cyanobacteria and methanogens embedded in Mars analogue minerals by the use of Raman spectroscopy. In Proceedings of the 2012 EGU General Assembly Conference, Vienna, Austria, 22–27 April 2012; Volume 14, p. 2334.
- Archer, D.B. Uncoupling of Methanogenesis from Growth of Methanosarcina barkeri by Phosphate Limitation. Appl. Environ. Microbiol. 1985, 50, 1233–1237. [Google Scholar] [PubMed]
- Liu, J.S.; Schill, N.; van Gulik, W.M.; Voisard, D.; Marison, I.W.; von Stockar, U. The coupling between catabolism and anabolism of Methanobacterium thermoautotrophicum in H2- and iron-limited continuous cultures. Enzym. Microb. Technol. 1999, 25, 784–794. [Google Scholar] [CrossRef]
- Fardeau, M.L.; Belaich, J.P. Energetics of the growth of Methanococcus thermolithotrophicus. Arch. Microbiol. 1986, 144, 381–385. [Google Scholar] [CrossRef]
- Tsao, J.H.; Kaneshiro, S.M.; Yu, S.S.; Clark, D.S. Continuous culture of Methanococcus jannaschii, an extremely thermophilic methanogen. Biotechnol. Bioeng. 1994, 43, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Mountfort, D.O.; Asher, R.A. Effect of inorganic sulfide on the growth and metabolism of Methanosarcina barkeri strain DM. Appl. Environ. Microbiol. 1979, 37, 670–675. [Google Scholar] [PubMed]
- Pennings, J.L.A.; Vermeij, P.; Poorter, L.M.I.D.; Keltjens, J.T.; Vogels, G.D. Adaptation of methane formation and enzyme contents during growth of Methanobacterium thermoautotrophicum (strain ΔH) in a fed-batch fermentor. Antonie Leeuwenhoek 2000, 77, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Ver Eecke, H.C.; Akerman, N.H.; Huber, J.A.; Butterfield, D.A.; Holden, J.F. Growth kinetics and energetics of a deep-sea hyperthermophilic methanogen under varying environmental conditions. Environ. Microbiol. Rep. 2013, 5, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Ver Eecke, H.C.V.; Butterfield, D.A.; Huber, J.A.; Lilley, M.D.; Olson, E.J.; Roe, K.K.; Evans, L.J.; Merkel, A.Y.; Cantin, H.V.; Holden, J.F. Hydrogen-limited growth of hyperthermophilic methanogens at deep-sea hydrothermal vents. Proc. Natl. Acad. Sci. USA 2012, 109, 13674–13679. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Nakamura, K.; Toki, T.; Tsunogai, U.; Miyazaki, M.; Miyazaki, J.; Hirayama, H.; Nakagawa, S.; Nunoura, T.; Horikoshi, K. Cell proliferation at 122 degrees C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proc. Natl. Acad. Sci. USA 2008, 105, 10949–10954. [Google Scholar] [CrossRef] [PubMed]
- Morozova, D.; Wagner, D. Stress response of methanogenic archaea from Siberian permafrost compared with methanogens from nonpermafrost habitats. FEMS Microbiol. Ecol. 2007, 61, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schönheit, P.; Moll, J.; Thauer, R.K. Growth parameters (Ks, μmax, Ys) of Methanobacterium thermoautotrophicum. Arch. Microbiol. 1980, 127, 59–65. [Google Scholar] [CrossRef]
- Lai, M.C.; Chen, S.C.; Shu, C.M.; Chiou, M.S.; Wang, C.C.; Chuang, M.J.; Hong, T.Y.; Liu, C.C.; Lai, L.J.; Hua, J.J. Methanocalculus taiwanensis sp. nov., isolated from an estuarine environment. Int. J. Syst. Evol. Microbiol. 2002, 52, 1799–1806. [Google Scholar] [CrossRef] [PubMed]
- The latest findings show, that there is the possibility of hydrothermal vents on icy moons like Europa (e.g., [215]) or Enceladus [202], which would widen the potential temperature range for microbes in the outer Solar System.
- Dong, X.; Chen, Z. Psychrotolerant methanogenic archaea: Diversity and cold adaptation mechanisms. Sci. China Life Sci. 2012, 55, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.S.; Cavicchioli, R. Cold-Adapted Enzymes. Annu. Rev. Biochem. 2006, 75, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Feller, G.; Gerday, C. Psychrophilic enzymes: Hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.; Cavicchioli, R.; Thomas, T. Thermodynamic activation properties of elongation factor 2 (EF-2) proteins from psychrotolerant and thermophilic Archaea. Extremophiles 2002, 6, 143–150. [Google Scholar] [PubMed]
- Thomas, T.; Cavicchioli, R. Effect of Temperature on Stability and Activity of Elongation Factor 2 Proteins from Antarctic and Thermophilic Methanogens. J. Bacteriol. 2000, 182, 1328–1332. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Kumar, N.; Cavicchioli, R. Effects of Ribosomes and Intracellular Solutes on Activities and Stabilities of Elongation Factor 2 Proteins from Psychrotolerant and Thermophilic Methanogens. J. Bacteriol. 2001, 183, 1974–1982. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Thomas, T.; Cavicchioli, R. Low temperature regulated DEAD-box RNA helicase from the antarctic archaeon, Methanococcoides burtonii. J. Mol. Biol. 2000, 297, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Liu, X.; Dong, X. Methanospirillum psychrodurum sp. nov., isolated from wetland soil. Int. J. Syst. Evol. Microbiol. 2014, 64, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Simankova, M.V.; Parshina, S.N.; Tourova, T.P.; Kolganova, T.V.; Zehnder, A.J.; Nozhevnikova, A.N. Methanosarcina lacustris sp. nov., a new psychrotolerant methanogenic archaeon from anoxic lake sediments. Syst. Appl. Microbiol. 2001, 24, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Jiang, N.; Liu, X.; Dong, X. Methanogenesis from Methanol at Low Temperatures by a Novel Psychrophilic Methanogen, “Methanolobus psychrophilus” sp. nov., Prevalent in Zoige Wetland of the Tibetan Plateau. Appl. Environ. Microbiol. 2008, 74, 6114–6120. [Google Scholar] [CrossRef] [PubMed]
- Chong, S.C.; Liu, Y.; Cummins, M.; Valentine, D.L.; Boone, D.R. Methanogenium marinum sp. nov., a H2-using methanogen from Skan Bay, Alaska, and kinetics of H2 utilization. Antonie Van Leeuwenhoek 2002, 81, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Zhilina, T.N.; Zavarzin, G.A. Methanohalobium evestigatus, n. gen., n. sp. The extremely halophilic methanogenic Archaebacterium. Dokl. Akad. Nauk SSSR 1987, 293, 464–468. [Google Scholar]
- Romesser, J.A.; Wolfe, R.S.; Mayer, F.; Spiess, E.; Walther-Mauruschat, A. Methanogenium, a new genus of marine methanogenic bacteria, and characterization of Methanogenium cariaci sp. nov. and Methanogenium marisnigri sp. nov. Arch. Microbiol. 1979, 121, 147–153. [Google Scholar] [CrossRef]
- Bräuer, S.L.; Cadillo-Quiroz, H.; Ward, R.J.; Yavitt, J.B.; Zinder, S.H. Methanoregula boonei gen. nov., sp. nov., an acidiphilic methanogen isolated from an acidic peat bog. Int. J. Syst. Evol. Microbiol. 2011, 61, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Maestrojuán, G.M.; Boone, D.R.; Xun, L.; Mah, R.A.; Zhang, L. Transfer of Methanogenium bourgense, Methanogenium marisnigri, Methanogenium olentangyi, and Methanogenium thermophilicum to the Genus Methanoculleus gen. nov., Emendation of Methanoculleus marisnigri and Methanogenium, and Description of New Strains of Methanoculleus bourgense and Methanoculleus marisnigri. Int. J. Syst. Bacteriol. 1990, 40, 117–122. [Google Scholar]
- Dianou, D.; Miyaki, T.; Asakawa, S.; Morii, H.; Nagaoka, K.; Oyaizu, H.; Matsumoto, S. Methanoculleus chikugoensis sp. nov., a novel methanogenic archaeon isolated from paddy field soil in Japan, and DNA-DNA hybridization among Methanoculleus species. Int. J. Syst. Evol. Microbiol. 2001, 51, 1663–1669. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Kendall, M.M.; Liu, Y.; Boone, D.R. Isolation and characterization of methylotrophic methanogens from anoxic marine sediments in Skan Bay, Alaska: Description of Methanococcoides alaskense sp. nov., and emended description of Methanosarcina baltica. Int. J. Syst. Evol. Microbiol. 2005, 55, 2531–2538. [Google Scholar] [CrossRef] [PubMed]
- Franzmann, P.D.; Springer, N.; Ludwig, W.; De Macario, E.C.; Rohde, M. A Methanogenic Archaeon from Ace Lake, Antarctica: Methanococcoides burtonii sp. nov. Syst. Appl. Microbiol. 1992, 15, 573–581. [Google Scholar] [CrossRef]
- Shimada, Y.; Fukuda, W.; Akada, Y.; Ishida, M.; Nakayama, J.; Imanaka, T.; Fujiwara, S. Property of cold inducible DEAD-box RNA helicase in hyperthermophilic archaea. Biochem. Biophys. Res. Commun. 2009, 389, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Saunders, N.F.W.; Thomas, T.; Curmi, P.M.G.; Mattick, J.S.; Kuczek, E.; Slade, R.; Davis, J.; Franzmann, P.D.; Boone, D.; Rusterholtz, K.; et al. Mechanisms of Thermal Adaptation Revealed From the Genomes of the Antarctic Archaea Methanogenium frigidum and Methanococcoides burtonii. Genome Res. 2003, 13, 1580–1588. [Google Scholar] [CrossRef] [PubMed]
- Gunnigle, E.; McCay, P.; Fuszard, M.; Botting, C.H.; Abram, F.; O’Flaherty, V. A Functional Approach to Uncover the Low-Temperature Adaptation Strategies of the Archaeon Methanosarcina barkeri. Appl. Environ. Microbiol. 2013, 79, 4210–4219. [Google Scholar] [CrossRef] [PubMed]
- Koga, Y. Thermal adaptation of the archaeal and bacterial lipid membranes. Archaea (Vanc. B.C.) 2012, 2012, 789652. [Google Scholar] [CrossRef] [PubMed]
- Nichols, D.S.; Miller, M.R.; Davies, N.W.; Goodchild, A.; Raftery, M.; Cavicchioli, R. Cold adaptation in the Antarctic Archaeon Methanococcoides burtonii involves membrane lipid unsaturation. J. Bacteriol. 2004, 186, 8508–8515. [Google Scholar] [CrossRef] [PubMed]
- Sprott, G.D.; Meloche, M.; Richards, J.C. Proportions of diether, macrocyclic diether, and tetraether lipids in Methanococcus jannaschii grown at different temperatures. J. Bacteriol. 1991, 173, 3907–3910. [Google Scholar] [PubMed]
- Rittmann, S.; Herwig, C. A comprehensive and quantitative review of dark fermentative biohydrogen production. Microb. Cell Fact. 2012, 11, 115. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, N.; Kitaura, S.; Mimura, A.; Takahara, Y. Cultivation of thermophilic methanogen KN-15 on H2-CO2 under pressurized conditions. J. Ferment. Bioeng. 1992, 73, 477–480. [Google Scholar] [CrossRef]
- Park, C.B.; Clark, D.S. Rupture of the Cell Envelope by Decompression of the Deep-Sea Methanogen Methanococcus jannaschii. Appl. Environ. Microbiol. 2002, 68, 1458–1463. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.F.; Shah, N.N.; Nelson, C.M.; Ludlow, J.M.; Clark, D.S. Pressure and Temperature Effects on Growth and Methane Production of the Extreme Thermophile Methanococcus jannaschii. Appl. Environ. Microbiol. 1988, 54, 3039–3042. [Google Scholar] [PubMed]
- Boonyaratanakornkit, B.; Córdova, J.; Park, C.B.; Clark, D.S. Pressure affects transcription profiles of Methanocaldococcus jannaschii despite the absence of barophilic growth under gas-transfer limitation. Environ. Microbiol. 2006, 8, 2031–2035. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, G.; Jaenicke, R.; Ludemann, H.D. High-Pressure Equipment for Growing Methanogenic Microorganisms on Gaseous Substrates at High Temperature. Appl. Environ. Microbiol. 1987, 53, 1876–1879. [Google Scholar] [PubMed]
- Mayumi, D.; Dolfing, J.; Sakata, S.; Maeda, H.; Miyagawa, Y.; Ikarashi, M.; Tamaki, H.; Takeuchi, M.; Nakatsu, C.H.; Kamagata, Y. Carbon dioxide concentration dictates alternative methanogenic pathways in oil reservoirs. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Huber, G.; Stetter, K.O. Sulfolobus metallicus, sp. nov., a Novel Strictly Chemolithoautotrophic Thermophilic Archaeal Species of Metal-Mobilizers. Syst. Appl. Microbiol. 1991, 14, 372–378. [Google Scholar] [CrossRef]
- Suzuki, T.; Iwasaki, T.; Uzawa, T.; Hara, K.; Nemoto, N.; Kon, T.; Ueki, T.; Yamagishi, A.; Oshima, T. Sulfolobus tokodaii sp. nov. (f. Sulfolobus sp. strain 7), a new member of the genus Sulfolobus isolated from Beppu Hot Springs, Japan. Extremophiles 2002, 6, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Dong, X.; Huang, L. Sulfolobus tengchongensis sp. nov., a novel thermoacidophilic archaeon isolated from a hot spring in Tengchong, China. Extremophiles 2003, 7, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Schleper, C.; Piihler, G.; Kuhlmorgen, B.; Zillig, W. Life at extremely low pH. Nature 1995, 375, 741–742. [Google Scholar] [CrossRef] [PubMed]
- Cadillo-Quiroz, H.; Yavitt, J.B.; Zinder, S.H. Methanosphaerula palustris gen. nov., sp. nov., a hydrogenotrophic methanogen isolated from a minerotrophic fen peatland. Int. J. Syst. Evol. Microbiol. 2009, 59, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Zhilina, T.N.; Zavarzina, D.G.; Kevbrin, V.V.; Kolganova, T.V. Methanocalculus natronophilus sp. nov., a new alkaliphilic hydrogenotrophic methanogenic archaeon from a soda lake, and proposal of the new family Methanocalculaceae. Microbiology 2013, 82, 698–706. [Google Scholar] [CrossRef]
- Rea, S.; Bowman, J.P.; Popovski, S.; Pimm, C.; Wright, A.D.G. Methanobrevibacter millerae sp. nov. and Methanobrevibacter olleyae sp. nov., methanogens from the ovine and bovine rumen that can utilize formate for growth. Int. J. Syst. Evol. Microbiol. 2007, 57, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Burggraf, S.; Fricke, H.; Neuner, A.; Kristjansson, J.; Rouvier, P.; Mandelco, L.; Woese, C.R.; Stetter, K.O. Methanococcus igneus sp. nov., a Novel Hyperthermophilic Methanogen from a Shallow Submarine Hydrothermal System. Syst. Appl. Microbiol. 1990, 13, 263–269. [Google Scholar] [CrossRef]
- Jones, W.J.; Paynter, M.J.B.; Gupta, R. Characterization of Methanococcus maripaludis sp. nov., a new methanogen isolated from salt marsh sediment. Arch. Microbiol. 1983, 135, 91–97. [Google Scholar] [CrossRef]
- Robertson, D.E.; Noll, D.; Roberts, M.F.; Menaia, J.A.; Boone, D.R. Detection of the osmoregulator betaine in methanogens. Appl. Environ. Microbiol. 1990, 56, 563–565. [Google Scholar] [PubMed]
- Proctor, L.M.; Lai, R.; Gunsalus, R.P. The methanogenic archaeon Methanosarcina thermophila TM-1 possesses a high-affinity glycine betaine transporter involved in osmotic adaptation. Appl. Environ. Microbiol. 1997, 63, 2252–2257. [Google Scholar] [PubMed]
- Sowers, K.R.; Boone, J.E.; Gunsalus, R.P. Disaggregation of Methanosarcina spp. and Growth as Single Cells at Elevated Osmolarity. Appl. Environ. Microbiol. 1993, 59, 3832–3839. [Google Scholar] [PubMed]
- Lai, M.C.; Sowers, K.R.; Robertson, D.E.; Roberts, M.F.; Gunsalus, R.P. Distribution of compatible solutes in the halophilic methanogenic archaebacteria. J. Bacteriol. 1991, 173, 5352–5358. [Google Scholar] [PubMed]
- Roeßler, M.; Pflüger, K.; Flach, H.; Lienard, T.; Gottschalk, G.; Müller, V. Identification of a Salt-Induced Primary Transporter for Glycine Betaine in the Methanogen Methanosarcina mazei Gö1. Appl. Environ. Microbiol. 2002, 68, 2133–2139. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.C.; Hong, T.Y.; Gunsalus, R.P. Glycine Betaine Transport in the Obligate Halophilic Archaeon Methanohalophilus portucalensis. J. Bacteriol. 2000, 182, 5020–5024. [Google Scholar] [CrossRef] [PubMed]
- Cockell, C.; Catling, D.; Wanda, L.; Snook, K.; Kepner, R.; Lee, P.; McKay, C. The Ultraviolet Environment of Mars: Biological Implications Past, Present, and Future. Icarus 2000, 146, 343–359. [Google Scholar] [CrossRef] [PubMed]
- Beatty, J.; Petersen, C.; Chaikin, A. (Eds.) New Solar System; Sky Pub.: Cambridge, MA, USA, 1999.
- Fendrihan, S.; Bérces, A.; Lammer, H.; Musso, M.; Rontó, G.; Polacsek, T.K.; Holzinger, A.; Kolb, C.; Stan-Lotter, H. Investigating the Effects of Simulated Martian Ultraviolet Radiation on Halococcus dombrowskii and Other Extremely Halophilic Archaebacteria. Astrobiology 2009, 9, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Ajon, M.; Fröls, S.; van Wolferen, M.; Stoecker, K.; Teichmann, D.; Driessen, A.J.M.; Grogan, D.W.; Albers, S.V.; Schleper, C. UV-inducible DNA exchange in hyperthermophilic archaea mediated by type IV pili. Mol. Microbiol. 2011, 82, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Fröls, S.; Gordon, P.M.K.; Panlilio, M.A.; Duggin, I.G.; Bell, S.D.; Sensen, C.W.; Schleper, C. Response of the Hyperthermophilic Archaeon Sulfolobus solfataricus to UV Damage. J. Bacteriol. 2007, 189, 8708–8718. [Google Scholar] [CrossRef] [PubMed]
- Fröls, S.; Ajon, M.; Wagner, M.; Teichmann, D.; Zolghadr, B.; Folea, M.; Boekema, E.J.; Driessen, A.J.M.; Schleper, C.; Albers, S.V. UV-inducible cellular aggregation of the hyperthermophilic archaeon Sulfolobus solfataricus is mediated by pili formation. Mol. Microbiol. 2008, 70, 938–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fröls, S.; White, M.F.; Schleper, C. Reactions to UV damage in the model archaeon Sulfolobus solfataricus. Biochem. Soc. Trans. 2009, 37, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Kral, T.; Shina, N. Sensitivity of methanogens to ultraviolet radiation under aerobic and anaerobic conditions. In Proceedings of the 2012 Astrobiology Science Conference, Atlanta, GA, USA, 16–20 April 2012.
- Sinha, N.; Kral, T.A. Methanogen Sensitivity to Ultraviolet Radiation: Implications for Life on Mars. Meteorit. Planet. Sci. Suppl. 2013, 76, 5071. [Google Scholar]
- Stan-Lotter, H.; Legat, A.; Fendrihan, S.; Leuko, S.; Gruber, C.; Radax, C.; Pfaffenhuemer, M.; Weidler, G.; Rittmann, S. Haloarchaeal survival over geological times and the detection of extraterrestrial halite—Implications for the search for life on Mars. In Proceedings of the Third European Workshop on Exo-Astrobiology, Madrid, Spain, 18–20 November 2003; ESA Publications Division: Noordwijk, The Netherlands, 2004; Volume 545, pp. 63–66. [Google Scholar]
- Radax, C.; Pfaffenhuemer, M.; Wieland, H.; Rittmann, S.; Leuko, S.; Weidler, G.; Gruber, C.; Stan-Lotter, H. Microbes in rock salt: How to find out what is in there. In Proceedings of the First European Workshop on Exo-Astrobiology, Graz, Austria, 16–19 September 2002; ESA Publications Division: Noordwijk, The Netherlands, 2002; Volume 518, pp. 485–486. [Google Scholar]
- Rittmann, S.; Legat, A.; Fendrihan, S.; Stan-Lotter, H. Viability and morphology of Halobacterium species following desiccation—Implications for contaminants on Mars. In Proceedings of the Third European Workshop on Exo-Astrobiology, Madrid, Spain, 18–20 November 2003; ESA Publications Division: Noordwijk, The Netherlands, 2004; Volume 545, pp. 275–276. [Google Scholar]
- Leuko, S.; Weidler, G.; Rittmann, S.; Stan-Lotter, H. LIVE/DEAD Kit: A powerful tool to detect haloarchaeal survival (and life?) in unknown environmental samples. In Proceedings of the Third European Workshop on Exo-Astrobiology, Madrid, Spain, 18–20 November 2003; ESA Publications Division: Noordwijk, The Netherlands, 2004; Volume 545, pp. 231–232. [Google Scholar]
- Liu, C.T.; Miyaki, T.; Aono, T.; Oyaizu, H. Evaluation of methanogenic strains and their ability to endure aeration and water stress. Curr. Microbiol. 2008, 56, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Schirmack, J.; Alawi, M.; Wagner, D. Influence of Martian regolith analogs on the activity and growth of methanogenic archaea, with special regard to long-term desiccation. Extrem. Microbiol. 2015, 210. [Google Scholar] [CrossRef] [PubMed]
- Solar Dynamics Observatory/Atmospheric Imaging Assembly. The Sun at 304 Angstroms. 2010. Available online: http://sdo.gsfc.nasa.gov/assets/img/browse/2010/08/19/20100819_003221_4096_0304.jpg (accessed on 13 November 2015). [Google Scholar]
- International Astronomical Union. Eight Planets and New Solar System Designations. 2006. Available online: http://apod.nasa.gov/apod/ap060828.html (accessed on 13 November 2015). [Google Scholar]
- Tate, K. Dwarf Planets in the Solar System. 2015. Available online: http://i.space.com/images/i/000/023/868/original/dwarf-planets-121120b-02.jpg?1353517196 (accessed on 13 November 2015).
- National Aeronautics and Space Administration. Moons of the Solar System. 2004. Available online: http://solarsystem.nasa.gov/multimedia/gallery/Many_Moons-browse.jpg (accessed on 13 November 2015). [Google Scholar]
- Archinal, B.A.; A’Hearn, M.F.; Bowell, E.; Conrad, A.; Consolmagno, G.J.; Courtin, R.; Fukushima, T.; Hestroffer, D.; Hilton, J.L.; Krasinsky, G.A.; et al. Report of the IAU Working Group on Cartographic Coordinates and Rotational Elements: 2009. Celest. Mech. Dyn. Astron. 2011, 109, 101–135. [Google Scholar] [CrossRef]
- Chamberlin, A. Planetary Satellite Physical Parameters. Available online: http://ssd.jpl.nasa.gov/?sat_phys_par (accessed on 13 November 2015).
- National Aeronautics and Space Administration. Ten Cool Things Seen in the First Year of LRO. 2010. Available online: http://www.nasa.gov/mission_pages/LRO/news/first-year.html (accessed on 13 November 2015). [Google Scholar]
- Schenk, P. Atlas of the Galilean Satellites; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Chamberlin, A. Planets and Pluto: Physical Characteristics. Available online: http://ssd.jpl.nasa.gov/?planet_phys_par (accessed on 13 November 2015).
- Spohn, T.; Breuer, D.; Johnson, T. (Eds.) Encyclopedia of the Solar System, 3rd ed.; Elsevier: Philadelphia, PA, USA, 2014.
- Thomas, P.C.; Parker, J.W.; McFadden, L.A.; Russell, C.T.; Stern, S.A.; Sykes, M.V.; Young, E.F. Differentiation of the asteroid Ceres as revealed by its shape. Nature 2005, 437, 224–226. [Google Scholar] [CrossRef] [PubMed]
- Delitsky, M.L.; Lane, A.L. Ice chemistry on the Galilean satellites. J. Geophys. Res. 1998, 103, 31391–31404. [Google Scholar] [CrossRef]
- Moore, J.M.; Chapman, C.R.; Bierhaus, E.B.; Greeley, R.; Chuang, F.C.; Klemaszewski, J.; Clark, R.N.; Dalton, J.B.; Hibbitts, C.A.; Schenk, P.M.; et al. Callisto. In Jupiter. The Planet, Satellites and Magnetosphere; Bagenal, F., Dowling, T.E., McKinnon, W.B., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 397–426. [Google Scholar]
- Jaumann, R.; Kirk, R.L.; Lorenz, R.D.; Lopes, R.M.C.; Stofan, E.; Turtle, E.P.; Keller, H.U.; Wood, C.A.; Sotin, C.; Soderblom, L.A.; et al. Geology and Surface Processes on Titan. In Titan from Cassini-Huygens; Brown, R.H., Lebreton, J.P., Waite, J.H., Eds.; Springer: Berlin, Germany, 2010; p. 75. [Google Scholar]
- Thomas, P.C. Sizes, shapes, and derived properties of the saturnian satellites after the Cassini nominal mission. Icarus 2010, 208, 395–401. [Google Scholar] [CrossRef]
- Spencer, J.R.; Pearl, J.C.; Segura, M.; Flasar, F.M.; Mamoutkine, A.; Romani, P.; Buratti, B.J.; Hendrix, A.R.; Spilker, L.J.; Lopes, R.M.C. Cassini Encounters Enceladus: Background and the Discovery of a South Polar Hot Spot. Science 2006, 311, 1401–1405. [Google Scholar] [CrossRef] [PubMed]
- Porco, C.C.; Helfenstein, P.; Thomas, P.C.; Ingersoll, A.P.; Wisdom, J.; West, R.; Neukum, G.; Denk, T.; Wagner, R.; Roatsch, T.; et al. Cassini Observes the Active South Pole of Enceladus. Science 2006, 311, 1393–1401. [Google Scholar] [CrossRef] [PubMed]
- Iess, L.; Stevenson, D.J.; Parisi, M.; Hemingway, D.; Jacobson, R.A.; Lunine, J.I.; Nimmo, F.; Armstrong, J.W.; Asmar, S.W.; Ducci, M.; et al. The Gravity Field and Interior Structure of Enceladus. Science 2014, 344, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Buie, M.W.; Grundy, W.M.; Young, E.F.; Young, L.A.; Stern, S.A. Orbits and Photometry of Pluto’s Satellites: Charon, S/2005 P1, and S/2005 P2. AJ 2006, 132, 290–298. [Google Scholar] [CrossRef]
- Stern, S.A.; Trafton, L.M. On the Atmospheres of Objects in the Kuiper Belt. In The Solar System Beyond Neptune; Barucci, M.A., Boehnhardt, H., Cruikshank, D.P., Morbidelli, A., Dotson, R., Eds.; University of Arizona Press: Tucson, AZ, USA, 2008; pp. 365–380. [Google Scholar]
- Vance, S.; Bouffard, M.; Choukroun, M.; Sotin, C. Ganymede’s internal structure including thermodynamics of magnesium sulfate oceans in contact with ice. Planet. Space Sci. 2014, 96, 62–70. [Google Scholar] [CrossRef]
- Lollar, B.S.; McCollom, T.M. Geochemistry: Biosignatures and abiotic constraints on early life. Nature 2006, 444, E18. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Marmolejo, A.; Segura, A.; Escobar-Briones, E. Abiotic Production of Methane in Terrestrial Planets. Astrobiology 2013, 13, 550–559. [Google Scholar] [CrossRef] [PubMed]
- White, W. Isotope Geochemistry; Wiley Works; Wiley: Hoboken, NJ, USA, 2015. [Google Scholar]
- Mahaffy, P.R.; Webster, C.R.; Atreya, S.K.; Franz, H.; Wong, M.; Conrad, P.G.; Harpold, D.; Jones, J.J.; Leshin, L.A.; Manning, H.; et al. Abundance and Isotopic Composition of Gases in the Martian Atmosphere from the Curiosity Rover. Science 2013, 341, 263–266. [Google Scholar] [PubMed]
- Hoekzema, N.; Gwinner, K.; Grieger, B.; Markiewicz, W.J.; Keller, H.; Hoffmann, H.; Meima, J.A.; Neukum, G. The Dust Scale Height of the Martian Atmosphere around Pavonis Mons from Hrsc Stereo Images. In AAS/Division for Planetary Sciences Meeting Abstracts; AAS/Division for Planetary Sciences Meeting Abstracts #38; American Astronomical Society: Washington, DC, USA, 2006; Volume 38, p. #60.30. [Google Scholar]
- Williams, D.R. Mars Fact Sheet. Available online: http://nssdc.gsfc.nasa.gov/planetary/factsheet/marsfact.html (accessed on 19 November 2015).
- Williams, D.R. Earth Fact Sheet. Available online: http://nssdc.gsfc.nasa.gov/planetary/factsheet/earthfact.html (accessed on 19 November 2015).
- Lowell, P. Mars and Its Canals; Macmillan And Company Limited: London, UK, 1911. [Google Scholar]
- Villanueva, G.L.; Mumma, M.J.; Novak, R.E.; Käufl, H.U.; Hartogh, P.; Encrenaz, T.; Tokunaga, A.; Khayat, A.; Smith, M.D. Strong water isotopic anomalies in the martian atmosphere: Probing current and ancient reservoirs. Science 2015, 348, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Ojha, L.; Wilhelm, M.B.; Murchie, S.L.; McEwen, A.S.; Wray, J.J.; Hanley, J.; Massé, M.; Chojnacki, M. Spectral evidence for hydrated salts in recurring slope lineae on Mars. Nature Geoscience 2015, 8, 829–832. [Google Scholar] [CrossRef]
- Feldman, W.C.; Pathare, A.; Maurice, S.; Prettyman, T.H.; Lawrence, D.J.; Milliken, R.E.; Travis, B.J. Mars Odyssey neutron data: 2. Search for buried excess water ice deposits at nonpolar latitudes on Mars. J. Geophys. Res. (Planets) 2011, 116, 11009. [Google Scholar] [CrossRef]
- Hand, E. Curiosity set to weigh in on Mars methane puzzle. Nature 2012. Available online: http://www.nature.com/news/curiosity-set-to-weigh-in-on-mars-methane-puzzle-1.11721 (accessed on 19 November 2015). [Google Scholar] [CrossRef]
- The potential CH4 signal was in fact coming from CO2 ice.
- Webster, C.R.; Mahaffy, P.R.; Atreya, S.K.; Flesch, G.J.; Farley, K.A.; Team, M.S. Low Upper Limit to Methane Abundance on Mars. Science 2013, 342, 355–357. [Google Scholar] [CrossRef] [PubMed]
- Zahnle, K.; Freedman, R.S.; Catling, D.C. Is there methane on Mars? Icarus 2011, 212, 493–503. [Google Scholar] [CrossRef]
- Fonti, S.; Marzo, G.A. Mapping the methane on Mars. A & A 2010, 512, A51. [Google Scholar]
- Minor sources are, e.g., UV radiation induced generation of CH4 from organic chemicals and released CH4 from CH4 hydrates (i.e., clathrates), that may be a record of past biological activity.
- Catling, D.C.; Cockell, C.S.; McKay, C.P. Ultraviolet Radiation on the Surface of Mars. In Proceedings of the Fifth International Conference on Mars, Pasadena, CA, USA, 18–23 July 1999; p. 6128.
- Gaidos, E.J.; Nealson, K.H.; Kirschvink, J.L. Life in Ice-Covered Oceans. Science 1999, 284, 1631–1633. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, W.B.; Pappalardo, R.T.; Khurana, K.K. Europa: Perspectives on an Ocean World. In Europa; Pappalardo, R.T., McKinnon, W.B., Khurana, K.K., Eds.; University of Arizona Space Science Series; University of Arizona Press: Tucson, AZ, USA, 2009; p. 697. [Google Scholar]
- Oren, A. The bioenergetic basis for the decrease in metabolic diversity at increasing salt concentrations: Implications for the functioning of salt lake ecosystems. Hydrobiologia 2001, 466, 61–72. [Google Scholar] [CrossRef]
- Hussmann, H.; Sohl, F.; Spohn, T. Subsurface oceans and deep interiors of medium-sized outer planet satellites and large trans-neptunian objects. Icarus 2006, 185, 258–273. [Google Scholar] [CrossRef]
- Turse, C.; Leitner, J.J.; Firneis, M.G.; Schulze-Makuch, D. Simulations of Prebiotic Chemistry under Post-Impact Conditions on Titan. Life 2013, 3, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Hand, K.P.; Chyba, C.F.; Priscu, J.C.; Carlson, R.W.; Nealson, K.H. Astrobiology and the Potential for Life on Europa. In Europa; Pappalardo, R.T., McKinnon, W.B., Khurana, K.K., Eds.; The University of Arizona Space Science Series; University of Arizona Press: Tucson, AZ, USA, 2009; p. 589. [Google Scholar]
- Chamberlin, A. Planetary Satellite Mean Orbital Parameters. Available online: http://ssd.jpl.nasa.gov/?sat_elem (accessed on 19 November 2015).
- Spencer, J. Planetary science: Enceladus with a grain of salt. Nature 2009, 459, 1067–1068. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, W.B. Effect of Enceladus’s rapid synchronous spin on interpretation of Cassini gravity. Geophys. Res. Lett. 2015, 42, 2137–2143. [Google Scholar] [CrossRef]
- Postberg, F.; Kempf, S.; Schmidt, J.; Brilliantov, N.; Beinsen, A.; Abel, B.; Buck, U.; Srama, R. Sodium salts in E-ring ice grains from an ocean below the surface of Enceladus. Nature 2009, 459, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Taubner, R.S.; Leitner, J.; Firneis, M.; Hitzenberger, R. Modelling the Interior Structure of Enceladus Based on the 2014’s Cassini Gravity Data. J. Orig. Life Evol. Biosph. 2015, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Schubert, G.; Anderson, J.D.; Travis, B.J.; Palguta, J. Enceladus: Present internal structure and differentiation by early and long-term radiogenic heating. Icarus 2007, 188, 345–355. [Google Scholar] [CrossRef]
- Postberg, F. Sodium salts in cryo-volcanic ice particles—Evidence for liquid water on Enceladus. In Proceedings of the CHARM Meeting, Leicester, UK, 29 May 2009; National Aeronautics and Space Administration: Washington, DC, USA.
- Postberg, F.; Schmidt, J.; Hillier, J.; Kempf, S.; Srama, R. A salt-water reservoir as the source of a compositionally stratified plume on Enceladus. Nature 2011, 474, 620–622. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Tajeddine, R.; Tiscareno, M.; Burns, J.; Joseph, J.; Loredo, T.; Helfenstein, P.; Porco, C. Enceladus’s measured physical libration requires a global subsurface ocean. Icarus 2016, 264, 37–47. [Google Scholar] [CrossRef]
- Bouquet, A.; Mousis, O.; Waite, J.H.; Picaud, S. Possible evidence for a methane source in Enceladus’ ocean. Geophys. Res. Lett. 2015, 42, 1334–1339. [Google Scholar] [CrossRef]
- Waite, J.H., Jr.; Lewis, W.S.; Magee, B.A.; Lunine, J.I.; McKinnon, W.B.; Glein, C.R.; Mousis, O.; Young, D.T.; Brockwell, T.; Westlake, J.; et al. Liquid water on Enceladus from observations of ammonia and 40Ar in the plume. Nature 2009, 460, 487–490. [Google Scholar] [CrossRef]
- Hsu, H.W.; Postberg, F.; Sekine, Y.; Shibuya, T.; Kempf, S.; Horányi, M.; Juhász, A.; Altobelli, N.; Suzuki, K.; Masaki, Y.; et al. Ongoing hydrothermal activities within Enceladus. Nature 2015, 519, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Howett, C.; Spencer, J.; Verbiscer, A. Enceladus’ enigmatic heat flow. In AAS/Division for Planetary Sciences Meeting Abstracts; American Astronomical Society: Washington, DC, USA, 2014; Volume 46, p. #405.02. [Google Scholar]
- Howett, C.J.A.; Spencer, J.R.; Pearl, J.; Segura, M. High heat flow from Enceladus’ south polar region measured using 10–600 cm-1 Cassini/CIRS data. J. Geophys. Res. (Planets) 2011, 116, 3003. [Google Scholar] [CrossRef]
- Hansen, C.J.; Shemansky, D.E.; Esposito, L.W.; Stewart, A.I.F.; Lewis, B.R.; Colwell, J.E.; Hendrix, A.R.; West, R.A.; Waite, J.H., Jr.; Teolis, B.; et al. The composition and structure of the Enceladus plume. Geophys. Res. Lett. 2011, 38, 11202. [Google Scholar] [CrossRef]
- Horányi, M.; Juhász, A.; Morfill, G.E. Large-scale structure of Saturn’s E-ring. Geophys. Res. Lett. 2008, 35, 4203. [Google Scholar] [CrossRef]
- Kempf, S.; Srama, R.; Moragas-Klostermeyer, G.; Postberg, F.; Horányi, M.; Schmidt, J.; Spahn, F. The Structure of Saturn’s E ring as seen by Cassini CDA. In Proceedings of the 2011 EPSC-DPS Joint Meeting, Nantes, France, 2–7 October 2011; p. 1643.
- Schubert, G.; Sohl, F.; Hussmann, H. Interior of Europa. In Europa; Pappalardo, R.T., McKinnon, W.B., Khurana, K.K., Eds.; The University of Arizona Space Science Series; University of Arizona Press: Tucson, AZ, USA, 2009; p. 353. [Google Scholar]
- Taubner, R.S.; Leitner, J.; Firneis, M.; Hitzenberger, R. Estimations on the Interior of Small Icy Bodies in the Solar System (EGU2014-7338). In Presented at the European Geosciences Union General Assembly, Vienna, Austria, 27 April–2 May 2014.
- Kivelson, M.G.; Khurana, K.K.; Russell, C.T.; Volwerk, M.; Walker, R.J.; Zimmer, C. Galileo Magnetometer Measurements: A Stronger Case for a Subsurface Ocean at Europa. Science 2000, 289, 1340–1343. [Google Scholar] [CrossRef] [PubMed]
- Carr, M.H.; Belton, M.J.S.; Chapman, C.R.; Davies, M.E.; Geissler, P.; Greenberg, R.; McEwen, A.S.; Tufts, B.R.; Greeley, R.; Sullivan, R.; et al. Evidence for a subsurface ocean on Europa. Nature 1998, 391, 363. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, R.; Geissler, P.; Hoppa, G.; Tufts, B.R. Tidal-Tectonic Processes and Their Implications for the Character of Europa’s Icy Crust. Rev. Geophys. 2002, 40, 1004. [Google Scholar] [CrossRef]
- Brown, M.E.; Hand, K.P. Salts and Radiation Products on the Surface of Europa. AJ 2013, 145, 110. [Google Scholar] [CrossRef]
- Roth, L.; Saur, J.; Retherford, K.D.; Strobel, D.F.; Feldman, P.D.; McGrath, M.A.; Nimmo, F. Transient Water Vapor at Europa’s South Pole. Science 2014, 343, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Zolotov, M.Y.; Kargel, J.S. On the Chemical Composition of Europa’s Icy Shell, Ocean, and Underlying Rocks. In Europa; Pappalardo, R.T., McKinnon, W.B., Khurana, K.K., Eds.; The University of Arizona Space Science Series; University of Arizona Press: Tucson, AZ, USA, 2009; p. 431. [Google Scholar]
- McCollom, T.M. Methanogenesis as a potential source of chemical energy for primary biomass production by autotrophic organisms in hydrothermal systems on Europa. J. Geophys. Res. Planets 1999, 104, 30729–30742. [Google Scholar] [CrossRef]
- Niemann, H.B.; Atreya, S.K.; Demick, J.E.; Gautier, D.; Haberman, J.A.; Harpold, D.N.; Kasprzak, W.T.; Lunine, J.I.; Owen, T.C.; Raulin, F. Composition of Titan’s lower atmosphere and simple surface volatiles as measured by the Cassini-Huygens probe gas chromatograph mass spectrometer experiment. J. Geophys. Res. (Planets) 2010, 115, 12006. [Google Scholar] [CrossRef]
- Raulin, F. Astrobiology and habitability of Titan. Space Sci. Rev. 2008, 135, 37–48. [Google Scholar] [CrossRef]
- Raulin, F.; McKay, C.; Lunine, J.; Owen, T. Titan’s Astrobiology. In Titan from Cassini-Huygens; Brown, R.H., Lebreton, J.P., Waite, J.H., Eds.; Springer Science & Business Media: Berlin, Germany, 2010; p. 215. [Google Scholar]
- Fulchignoni, M.; Ferri, F.; Angrilli, F.; Ball, A.J.; Bar-Nun, A.; Barucci, M.A.; Bettanini, C.; Bianchini, G.; Borucki, W.; Colombatti, G.; et al. In situ measurements of the physical characteristics of Titan’s environment. Nature 2005, 438, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.W.; Ferris, J.P. Chemical Evolution on Titan: Comparisons to the Prebiotic Earth. In Planetary and Interstellar Processes Relevant to the Origins of Life; Whittet, D.C.B., Ed.; Springer Science & Business Media: Berlin, Germany, 1997; p. 225. [Google Scholar]
- Balucani, N.; Cartechini, L.; Bergeat, A.; Casavecchia, P.; Volpi, G.G. Laboratory studies on the formation of CN containing molecules in the atmosphere of Titan and prebiotic Earth. In Proceedings of the First European Workshop on Exo-/Astro-Biology, Frascati, Italy, 21–23 May 2001; Ehrenfreund, P., Angerer, O., Battrick, B., Eds.; ESA Special Publication: Noordwijk, The Netherlands, 2001; Volume 496, pp. 159–162. [Google Scholar]
- Neish, C.D.; Somogyi, Á.; Imanaka, H.; Lunine, J.I.; Smith, M.A. Rate Measurements of the Hydrolysis of Complex Organic Macromolecules in Cold Aqueous Solutions: Implications for Prebiotic Chemistry on the Early Earth and Titan. Astrobiology 2008, 8, 273–287. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Smith, M.A. Identification of nitrogenous organic species in Titan aerosols analogs: Implication for prebiotic chemistry on Titan and early Earth. Icarus 2014, 238, 86–92. [Google Scholar] [CrossRef]
- Sotin, C.; Jaumann, R.; Buratti, B.J.; Brown, R.H.; Clark, R.N.; Soderblom, L.A.; Baines, K.H.; Bellucci, G.; Bibring, J.P.; Capaccioni, F.; et al. Release of volatiles from a possible cryovolcano from near-infrared imaging of Titan. Nature 2005, 435, 786–789. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, D.P.; Lorenz, R.D.; Lunine, J.I. Numerical calculations of the longevity of impact oases on Titan. Icarus 2005, 173, 243–253. [Google Scholar] [CrossRef]
- Iess, L.; Jacobson, R.A.; Ducci, M.; Stevenson, D.J.; Lunine, J.I.; Armstrong, J.W.; Asmar, S.W.; Racioppa, P.; Rappaport, N.J.; Tortora, P. The Tides of Titan. Science 2012, 337, 457–459. [Google Scholar] [CrossRef] [PubMed]
- Kerr, R.A. Cassini Spies an Ocean Inside Saturn’s Icy, Gassy Moon Titan. Science 2012, 336, 1629. [Google Scholar] [CrossRef] [PubMed]
- Sotin, C.; Mitri, G.; Rappaport, N.; Schubert, G.; Stevenson, D. Titan’s Interior Structure. In Titan from Cassini-Huygens; Brown, R.H., Lebreton, J.P., Waite, J.H., Eds.; Springer Science & Business Media: Berlin, Germany, 2010; p. 61. [Google Scholar]
- Buratti, B.; Faulk, S.; Mosher, J.; Baines, K.; Brown, R.; Clark, R.; Nicholson, P. Search for and limits on plume activity on Mimas, Tethys, and Dione with the Cassini Visual Infrared Mapping Spectrometer (VIMS). Icarus 2011, 214, 534–540. [Google Scholar] [CrossRef]
- There is an atmosphere indicating some material outflowing from the surface both at Dione and Rhea (J.H. Waite, personal communication, July, 2015).
- Tajeddine, R.; Rambaux, N.; Lainey, V.; Charnoz, S.; Richard, A.; Rivoldini, A.; Noyelles, B. Constraints on Mimas’ interior from Cassini ISS libration measurements. Science 2014, 346, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Gaeman, J.; Hier-Majumder, S.; Roberts, J.H. Sustainability of a subsurface ocean within Triton’s interior. Icarus 2012, 220, 339–347. [Google Scholar] [CrossRef]
- Soderblom, L.A.; Becker, T.L.; Kieffer, S.W.; Brown, R.H.; Hansen, C.J.; Johnson, T.V.; Kirk, R.L.; Shoemaker, E.M.; Cook, A.F. Triton’s geyser-like plumes—Discovery and basic characterization. Science 1990, 250, 410–415. [Google Scholar] [CrossRef] [PubMed]
- International Astronomical Union (Ed.) RESOLUTION B5—Definition of a Planet in the Solar System; International Astronomical Union: Prague, Czech Republic, 2006.
- Küppers, M.; O’Rourke, L.; Bockelée-Morvan, D.; Zakharov, V.; Lee, S.; von Allmen, P.; Carry, B.; Teyssier, D.; Marston, A.; Müller, T.; et al. Localized sources of water vapour on the dwarf planet (1)Ceres. Nature 2014, 505, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Witze, A. Mystery haze appears above Ceres’s bright spots. Nat. NEWS. 21 July 2015. Available online: http://www.nature.com/news/mystery-haze-appears-above-ceres-s-bright-spots-1.18032 (accessed on 19 November 2015).
- Robuchon, G.; Nimmo, F. Thermal evolution of Pluto and implications for surface tectonics and a subsurface ocean. Icarus 2011, 216, 426–439. [Google Scholar] [CrossRef]
- Rhoden, A.R.; Henning, W.; Hurford, T.A.; Hamilton, D.P. The interior and orbital evolution of Charon as preserved in its geologic record. Icarus 2015, 246, 11–20. [Google Scholar] [CrossRef]
- Large TNOs show a higher amount of rocky material, especially Quaoar with a mean density of 4200±1300 kg m-3 [270].
- Allen, C.C.; Morris, R.V.; Lindstrom, D.J.; Lindstrom, M.M.; Lockwood, J.P. JSC Mars-1—Martian regolith simulant. In Lunar and Planetary Institute Technical Report, Proceedings of the Lunar and Planetary Science Conference, Houston, TX, USA, 17–21 March 1997; Volume 28, p. 27.
- According to the reaction Fe0 + 2H+ → Fe2+ + H2, zero-valent iron could serve as reactant to produce H2 [67].
- Djordjevic, S.; Mickol, R.L.; Kral, T.A. Simulating Martian Conditions: Methanogen Survivability during Freeze-Thaw Cycles. In Proceedings of the Lunar and Planetary Science Conference, Woodlands, TX, USA, 17–21 March 2014; Volume 45, p. 2539.
- Mickol, R.L.; Kral, T.A.; Laird, S.K. Mesophile Methanogen Survival Under Freeze/Thaw Cycles. In Lunar and Planetary Institute Technical Report, Proceedings of the Lunar and Planetary Science Conference, Woodlands, TX, USA, 17–21 March 2014; Volume 45, p. 1603.
- Methanosarcina spec. SMA-16, SMA-23, M. soligelidi, Methanobacterium spec. MC-20, M. barkeri, and M. frigidum.
- McKay, C.P.; Porco, C.C.; Altheide, T.; Davis, W.L.; Kral, T.A. The Possible Origin and Persistence of Life on Enceladus and Detection of Biomarkers in the Plume. Astrobiology 2008, 8, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Found in the Columbia River basalts and basalts in the Twin Falls area of Idaho, respectively [246].
- Stevens, T.O.; McKinley, J.P. Lithoautotrophic Microbial Ecosystems in Deep Basalt Aquifers. Science 1995, 270, 450–455. [Google Scholar] [CrossRef]
- Chapelle, F.H.; O’Neill, K.; Bradley, P.M.; Methé, B.A.; Ciufo, S.A.; Knobel, L.L.; Lovley, D.R. A hydrogen-based subsurface microbial community dominated by methanogens. Nature 2002, 415, 312–315. [Google Scholar] [CrossRef] [PubMed]
- Found in the 3-to 4-km-deep fracture in the 2.7-billion-year-old Ventersdorp Supergroup metabasalt [251].
- Lin, L.H.; Wang, P.L.; Rumble, D.; Lippmann-Pipke, J.; Boice, E.; Pratt, L.M.; Sherwood Lollar, B.; Brodie, E.L.; Hazen, T.C.; Andersen, G.L.; et al. Long-Term Sustainability of a High-Energy, Low-Diversity Crustal Biome. Science 2006, 314, 479–482. [Google Scholar] [CrossRef] [PubMed]
- McKay, C.P.; Khare, B.N.; Amin, R.; Klasson, M.; Kral, T.A. Possible sources for methane and C2-C5 organics in the plume of Enceladus. Planet. Space Sci. 2012, 71, 73–79. [Google Scholar] [CrossRef]
- Parashar, S.; Kral, T.A. Possibility of Methanogens on Enceladus. LPI Contrib. 2010, 1538, 5044. [Google Scholar]
- Taubner, R.S.; Rittmann, S.; Leitner, J.; Schleper, C.; Firneis, M.; Hitzenberger, R. Assessing the feasibility to cultivate methanogens under Enceladus-like conditions reservoir. Presented at the 14th European Workshop on Astrobiology. In Presented at the 14th European Workshop on Astrobiology, Edinburgh, UK, 13–16 October 2014.
- Preston, L.J.; Dartnell, L.R. Planetary habitability: Lessons learned from terrestrial analogues. Int. J. Astrobiol. 2014, 13, 81–98. [Google Scholar] [CrossRef]
- Morris, R.V.; Blake, D.F.; Bish, D.; Ming, D.W.; Agresti, D.G.; Treiman, A.H.; Steele, A.; Amundsen, H.E.F.; Amase Team. A Terrestrial Analogue from Spitsbergen (Svalbard, Norway) for the Comanche Carbonate at Gusev Crater, Mars. In Lunar and Planetary Institute Technical Report, Proceedings of the Lunar and Planetary Science Conference, Woodlands, TX, USA, 7–11 March 2011; Volume 42, p. 1699.
- Gómez-Gómez, F.; Rodriguez-Manfredi, J.A.; Perez, L.; Prieto-Ballesteros, O.; Amils, R.; Gomez-Elvira, J. Martian Habitability Studies in Two Field Earth Analogues: The Permafrost in the Imuruk Lake Basaltic Field (Alaska) and the Atacama Desert. In Proceedings of the 38th COSPAR Scientific Assembly, Bremen, Germany, 15–18 July 2010; Volume 38, p. 3295.
- Shtarkman, Y.M.; Koçer, Z.A.; Edgar, R.; Veerapaneni, R.S.; D’Elia, T.; Morris, P.F.; Rogers, S.O. Subglacial Lake Vostok (Antarctica) Accretion Ice Contains a Diverse Set of Sequences from Aquatic, Marine and Sediment-Inhabiting Bacteria and Eukarya. PLoS ONE 2013, 8, e67221. [Google Scholar] [CrossRef] [PubMed]
- Patil, U.; Muskan, K. Essentials of Biotechnology; I.K. International Publishing House Pvt. Limited: New Deli, India, 2009. [Google Scholar]
- Fuchs, G.; Eitinger, T.; Schlegel, H. Allgemeine Mikrobiologie, 8th ed.; Thieme Georg Verlag: Stuttgart, Germany, 2007. [Google Scholar]
- Krajete, A.; Herwig, C.; Rittmann, S.; Seifert, A.; Bernacchi, S. Method and System for Producing Methane Using Methanogenic Microorganisms and Applying Specific Nitrogen Concentrations in the Liquid Phase. Patent WO2014128300 A1, 28 August 2014. [Google Scholar]
- Plaut, J.J.; Barabash, S.; Bruzzone, L.; Dougherty, M.; Erd, C.; Fletcher, L.; Gladstone, R.; Grasset, O.; Gurvits, L.; Hartogh, P.; et al. Jupiter Icy Moons Explorer (JUICE): Science Objectives, Mission and Instruments. In Proceedings of the Lunar and Planetary Science Conference, Woodlands, TX, USA, 17–21 March 2014; Volume 45, p. 2717.
- Grasset, O.; Dougherty, M.K.; Coustenis, A.; Bunce, E.J.; Erd, C.; Titov, D.; Blanc, M.; Coates, A.; Drossart, P.; Fletcher, L.N.; et al. JUpiter ICy moons Explorer (JUICE): An ESA mission to orbit Ganymede and to characterise the Jupiter system. Planet. Space Sci. 2013, 78, 1–21. [Google Scholar] [CrossRef]
- European Space Agency/Science and Robotic Exploration. JUICE Definition Study Report (Red Book); Technical Report 1.0; ESA: Paris, France, 2014. [Google Scholar]
- National Aeronautics and Space Administration. VALKYRIE: Phase 2. 2015. Available online: https://astrobiology.nasa.gov/astep/projects/nra/nnh11zda001n-astep/valkyrie-phase-2/ (accessed on 19 November 2015). [Google Scholar]
- Karl, D.M.; Beversdorf, L.; Björkman, K.; Church, M.J.; Martinez, A.; Delong, E.F. Aerobic production of methane in the sea. Nat. Geosci. 2008, 1, 473–478. [Google Scholar] [CrossRef]
- Metcalf, W.W.; Griffin, B.M.; Cicchillo, R.M.; Gao, J.; Janga, S.C.; Cooke, H.A.; Circello, B.T.; Evans, B.S.; Martens-Habbena, W.; Stahl, D.A.; et al. Synthesis of Methylphosphonic Acid by Marine Microbes: A Source for Methane in the Aerobic Ocean. Science 2012, 337, 1104–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carini, P.; White, A.; Campbell, E.; Giovannoni, S. Methane production by phosphate-starved SAR11 chemoheterotrophic marine bacteria. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Lecar, M.; Podolak, M.; Sasselov, D.; Chiang, E. On the Location of the Snow Line in a Protoplanetary Disk. ApJ 2006, 640, 1115–1118. [Google Scholar] [CrossRef]
- Fraser, W.C.; Brown, M.E. Quaoar: A Rock in the Kuiper Belt. ApJ 2010, 714, 1547–1550. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taubner, R.-S.; Schleper, C.; Firneis, M.G.; Rittmann, S.K.-M.R. Assessing the Ecophysiology of Methanogens in the Context of Recent Astrobiological and Planetological Studies. Life 2015, 5, 1652-1686. https://doi.org/10.3390/life5041652
Taubner R-S, Schleper C, Firneis MG, Rittmann SK-MR. Assessing the Ecophysiology of Methanogens in the Context of Recent Astrobiological and Planetological Studies. Life. 2015; 5(4):1652-1686. https://doi.org/10.3390/life5041652
Chicago/Turabian StyleTaubner, Ruth-Sophie, Christa Schleper, Maria G. Firneis, and Simon K.-M. R. Rittmann. 2015. "Assessing the Ecophysiology of Methanogens in the Context of Recent Astrobiological and Planetological Studies" Life 5, no. 4: 1652-1686. https://doi.org/10.3390/life5041652