Mexican Microalgae Biodiversity and State-Of-The-Art Extraction Strategies to Meet Sustainable Circular Economy Challenges: High-Value Compounds and Their Applied Perspectives

 ,
,  , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Mexican Microalgae Biodiversity
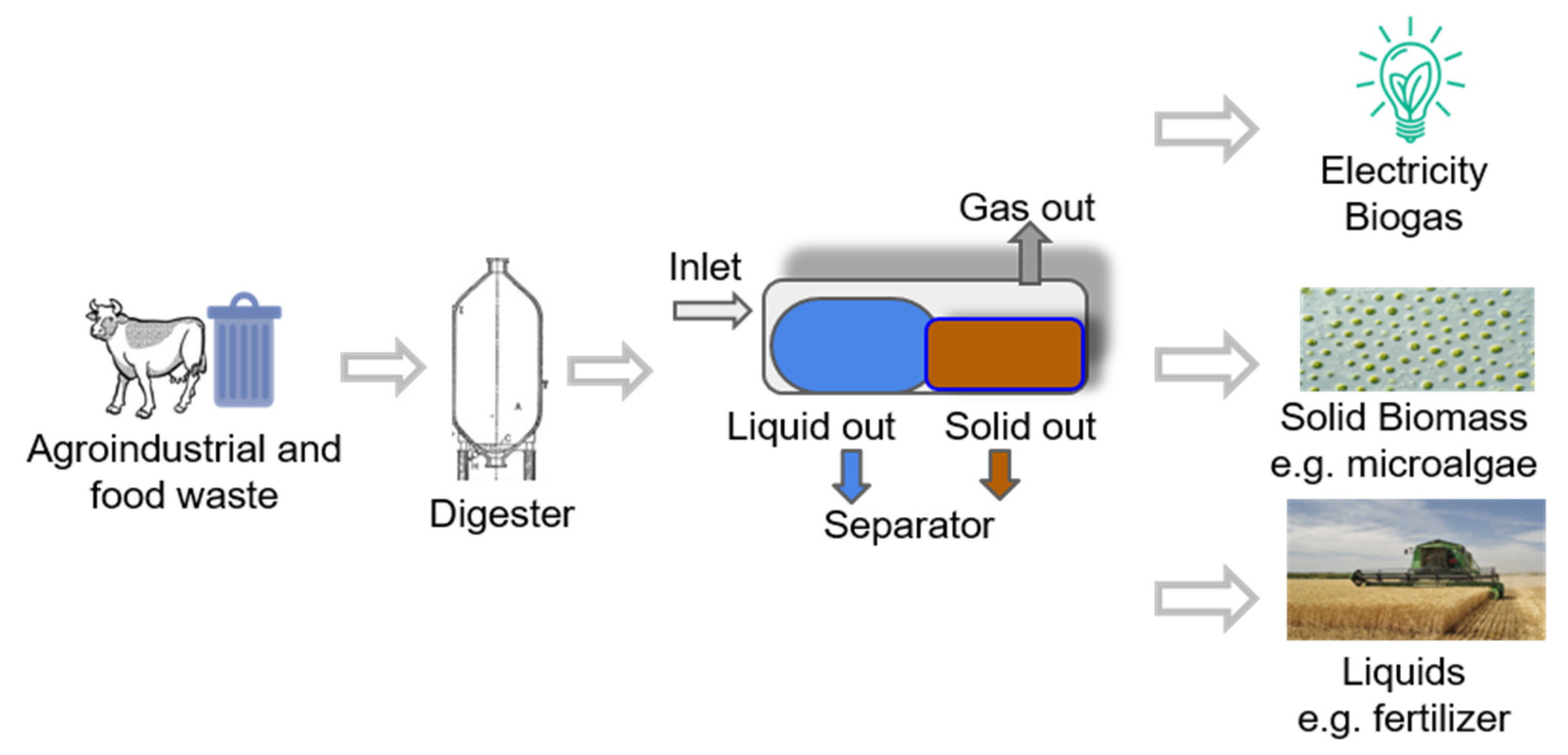
3. State-Of-The-Art of Extraction Methods
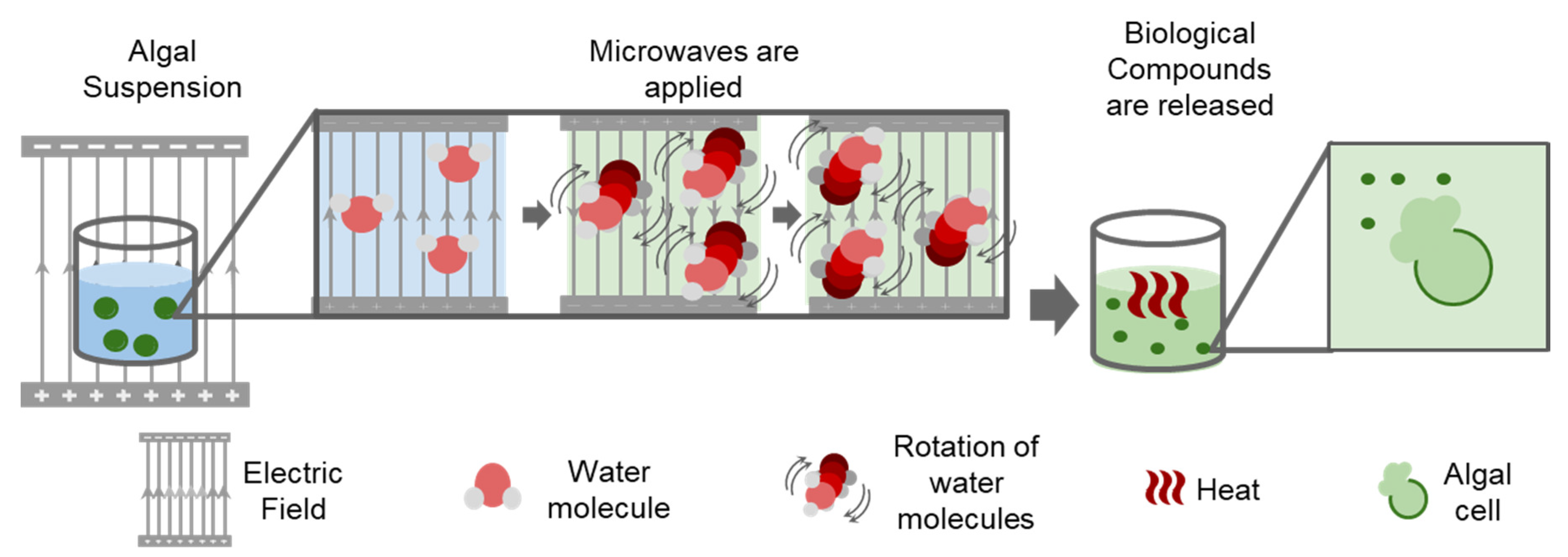
3.1. Microwave-Based Extraction
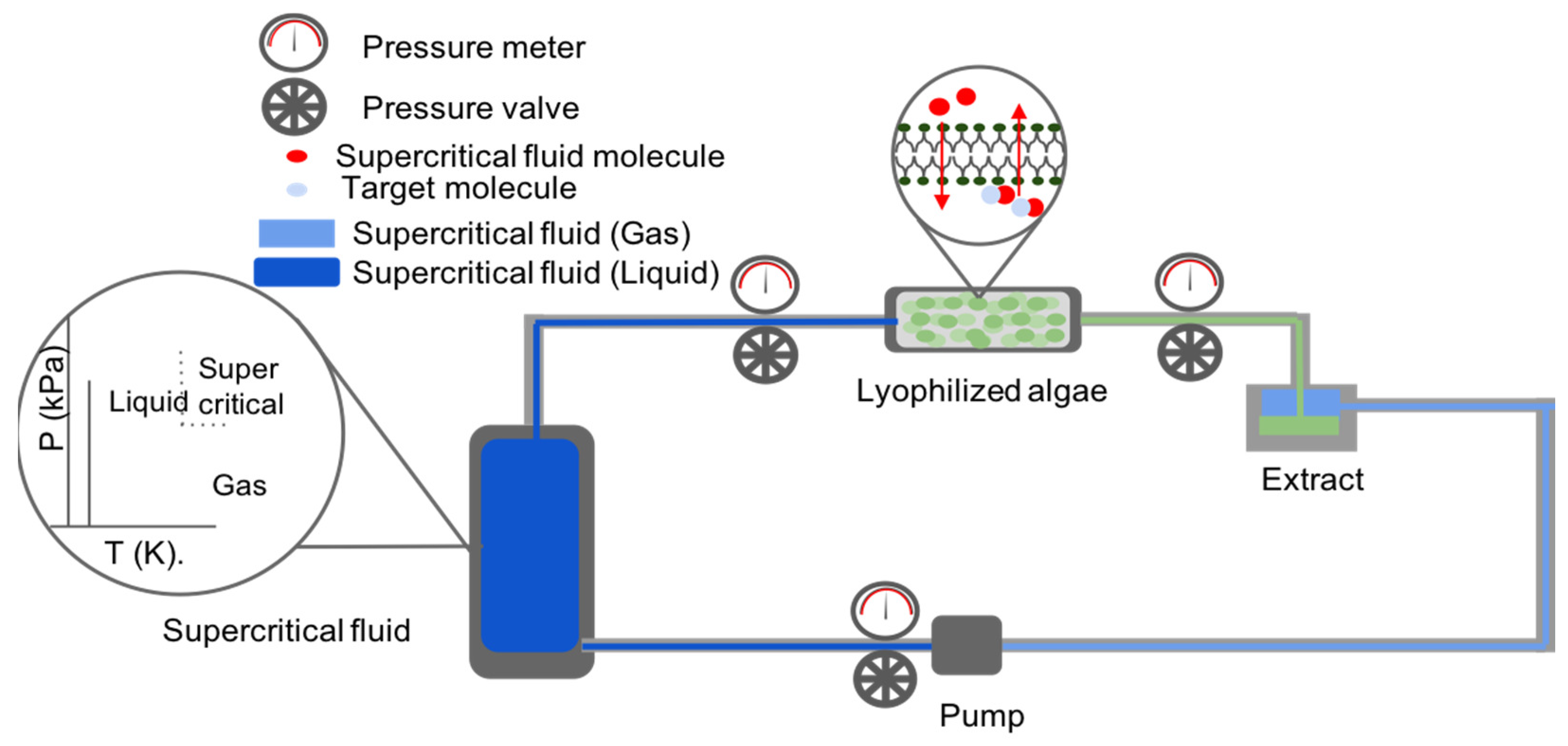
3.2. Supercritical Fluid-Based Extraction
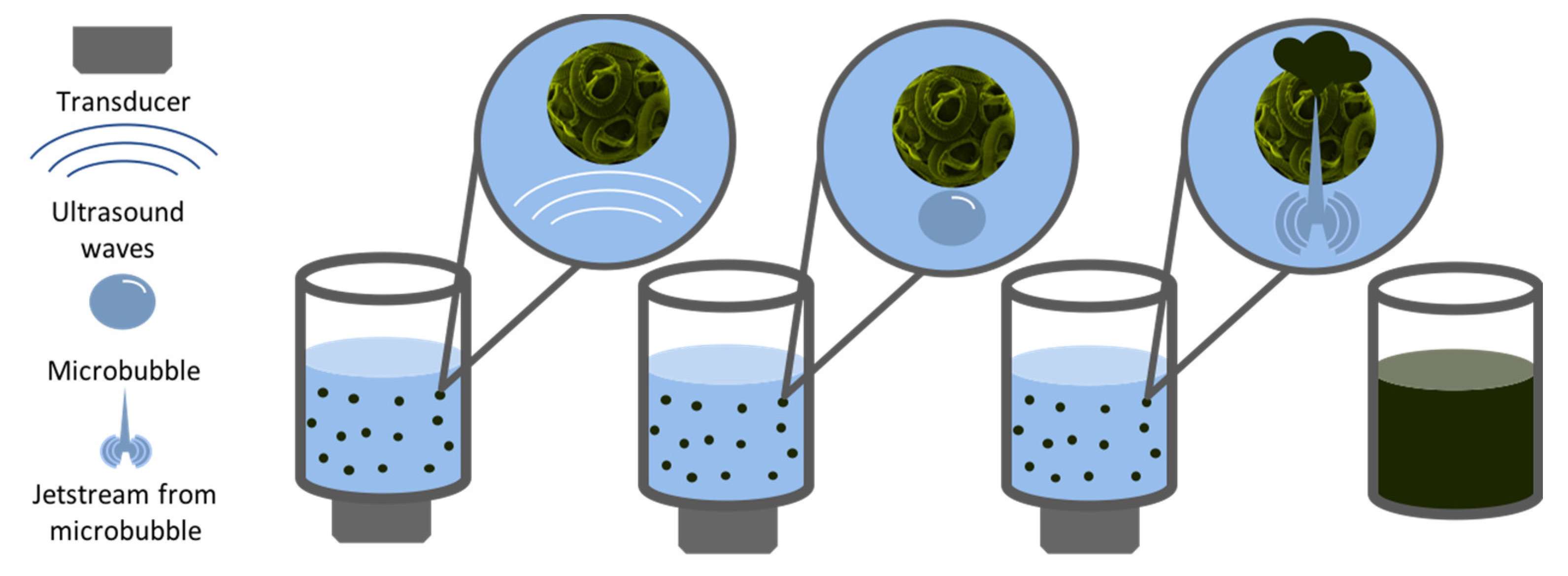
3.3. Ultrasound-Based Extraction
4. Current Applications
4.1. Pigments - Phycobilins, Lutein, and Carotenoids
4.2. Nutraceutical Potentialities
4.3. Bioactive Compounds
4.4. Bioremediation Potentialities
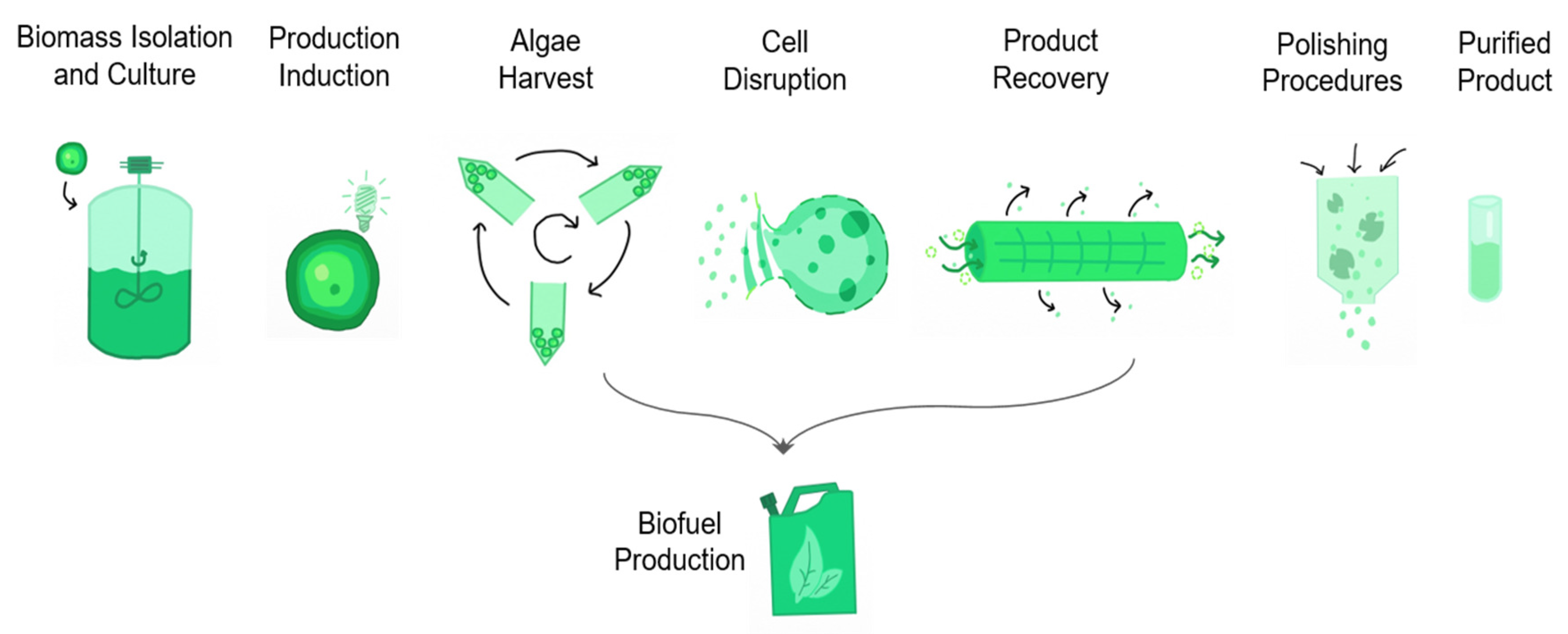
4.5. Bio-Fuels
4.5.1. Photosynthetically Production of Hydrogen
4.5.2. Biodiesel/Bioethanol
4.6. Antioxidants
4.7. Phycotoxins
5. Opportunities for Improvement
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Centella, M.H.; Arévalo-Gallegos, A.; Parra-Saldivar, R.; Iqbal, H.M. Marine-derived bioactive compounds for value-added applications in bio-and non-bio sectors. J. Clean. Prod. 2017, 168, 1559–1565. [Google Scholar] [CrossRef]
- Posten, C. Microalgae Biotechnology; Chen, S.F., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Kasting, J.F.; Siefert, J.L. Life and the evolution of Earth’s atmosphere. Science 2002, 296, 1066–1068. [Google Scholar] [CrossRef] [PubMed]
- Arbib, Z.; Ruiz, J.; Álvarez-Díaz, P.; Garrido-Perez, C.; Perales, J.A. Capability of different microalgae species for phytoremediation processes: Wastewater tertiary treatment, CO2 bio-fixation and low cost biofuels production. Water Res. 2014, 49, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Lei, A.P.; Hu, Z.L.; Wong, Y.S.; Tam, N.F.Y. Removal of fluoranthene and pyrene by different microalgal species. Bioresour. Technol. 2007, 98, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Mennaa, F.Z.; Arbib, Z.; Perales, J.A. Urban wastewater treatment by seven species of microalgae and an algal bloom: Biomass production, N and P removal kinetics and harvestability. Water Res. 2015, 83, 42–51. [Google Scholar] [CrossRef]
- Olaizola, M. Commercial development of microalgal biotechnology: From the test tube to the marketplace. Biomol. Eng. 2003, 20, 459–466. [Google Scholar] [CrossRef]
- Farrar, W.V. Tecuitlatl; a glimpse of Aztec food technology. Nature 1966, 211, 341–342. [Google Scholar] [CrossRef]
- Burlew, J.S. Algal Culture. From Laboratory to Pilot Plant; Carnegie Institution of Washington Publication: Washington, DC, USA, 1953; Volume 600. [Google Scholar]
- Belay, A.; Gershwin, M.E. Spirulina (Arthrospira). In Spirulina in Human Nutrition and Health; CRC Press: Boca Raton, FL, USA, 2007; pp. 11–35. [Google Scholar]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Ben-Amotz, A. Industrial production of microalgal cell-mass and secondary products-major industrial species. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2004; Volume 273. [Google Scholar]
- Lamers, P.P.; Janssen, M.; De Vos, R.C.; Bino, R.J.; Wijffels, R.H. Exploring and exploiting carotenoid accumulation in Dunaliella salina for cell-factory applications. Trends Biotechnol. 2008, 26, 631–638. [Google Scholar] [CrossRef]
- Jorquera, O.; Kiperstok, A.; Sales, E.A.; Embirucu, M.; Ghirardi, M.L. Comparative energy life-cycle analyses of microalgal biomass production in open ponds and photobioreactors. Bioresour. Technol. 2010, 101, 1406–1413. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853. [Google Scholar] [CrossRef]
- Oliva-Martínez, M.G.; Godínez-Ortega, J.L.; Zuñiga-Ramos, C.A. Biodiversidad del fitoplancton de aguas continentales en México. Rev. Mex. Biodivers. 2014, 85, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Muciño-Márquez, R.E.; Figueroa-Torres, M.G.; Aguirre-León, A. Cianofitas de los sistemas fluvio-lagunares Pom-Atasta y Palizada del Este, adyacentes a la Laguna de Términos, Campeche, México. Polibotánica 2015, 39, 49–78. [Google Scholar] [CrossRef]
- Jiménez-Valera, S.; del Pilar Sánchez-Saavedra, M. Growth and fatty acid profiles of microalgae species isolated from the Baja California Peninsula, México. Lat. Am. J. Aquat. Res. 2016, 44, 689–702. [Google Scholar] [CrossRef]
- Dávila-Camacho, C.A.; Galaviz-Villa, I.; Lango-Reynoso, F.; Castañeda-Chávez, M.D.R.; Quiroga-Brahms, C.; Montoya-Mendoza, J. Cultivation of native fish in Mexico: Cases of success. Rev. Aquac. 2018. [Google Scholar] [CrossRef]
- Rodríguez-Palacio, M.C.; Crisóstomo-Vázquez, L.; Álvarez-Hernández, S.; Lozano-Ramírez, C. Strains of toxic and harmful microalgae, from waste water, marine, brackish and fresh water. Food Addit. Contam. Part A 2012, 29, 304–313. [Google Scholar] [CrossRef]
- Ciferri, O.; Tiboni, O. The biochemistry and industrial potential of Spirulina. Annu. Rev. Microbiol. 1985, 39, 503–526. [Google Scholar] [CrossRef]
- Pacheco-Vega, J.M.; Cadena-Roa, M.A.; Ascencio, F.; Rangel-Dávalos, C.; Rojas-Contreras, M. Assessment of endemic microalgae as potential food for Artemia franciscana culture. Lat. Am. J. Aquat. Res. 2015, 43, 23–32. [Google Scholar] [CrossRef]
- Ramírez-Moreno, L.; Olvera-Ramírez, R. Uso tradicional y actual de Spirulina sp. (Arthrospira sp.). Interciencia 2006, 31, 657–663. [Google Scholar]
- Cea-Barcia, G.; Buitrón, G.; Moreno, G.; Kumar, G. A cost-effective strategy for the bio-prospecting of mixed microalgae with high carbohydrate content: Diversity fluctuations in different growth media. Bioresour. Technol. 2014, 163, 370–373. [Google Scholar] [CrossRef]
- Alcocer, J.; Hammer, U.T. Saline lake ecosystems of Mexico. Aquat. Ecosyst. Health Manag. 1998, 1, 291–315. [Google Scholar] [CrossRef]
- Toledo, G.; Bashan, Y.; Soeldner, A. Cyanobacteria and black mangroves in Northwestern Mexico: Colonization, and diurnal and seasonal nitrogen fixation on aerial roots. Can. J. Microbiol. 1995, 41, 999–1011. [Google Scholar] [CrossRef]
- Reyna-Martinez, R.; Gomez-Flores, R.; López-Chuken, U.; Quintanilla-Licea, R.; Caballero-Hernandez, D.; Rodríguez-Padilla, C.; Beltrán-Rocha, J.C.; Tamez-Guerra, P. Antitumor activity of Chlorella sorokiniana and Scenedesmus sp. microalgae native of Nuevo León State, México. PeerJ 2018, 6, e4358. [Google Scholar] [CrossRef]
- Poot-Delgado, C.A.; Okolodkov, Y.B.; Aké-Castillo, J.A. Potentially harmful cyanobacteria in oyster banks of Términos lagoon, southeastern Gulf of Mexico. Acta Biológica Colomb. 2018, 23, 51–58. [Google Scholar] [CrossRef] [Green Version]
- León-Tejera, H.; Montejano, G. Dermocarpella (Cyanoprokaryota/Cyanophyceae/Cyanobacteria) from the pacific coast of Mexico. Cryptogam. Algol. 2000, 21, 259–272. [Google Scholar] [CrossRef]
- Valadez, C.F.; Carmona, J.; Cantoral, U.E. Algas de ambientes lóticos en el estado de Morelos, México. An. Inst. Biol. Ser. Bot. 1996, 67, 227–282. [Google Scholar]
- Komolafe, O.; Orta, S.B.V.; Monje-Ramirez, I.; Noguez, I.Y.; Harvey, A.P.; Ledesma, M.T.O. Biodiesel production from indigenous microalgae grown in wastewater. Bioresour. Technol. 2014, 154, 297–304. [Google Scholar] [CrossRef]
- Toledo-Cervantes, A.; Morales, M.; Novelo, E.; Revah, S. Carbon dioxide fixation and lipid storage by Scenedesmus obtusiusculus. Bioresour. Technol. 2013, 130, 652–658. [Google Scholar] [CrossRef]
- Arzate-Cárdenas, M.A.; Olvera-Ramírez, R.; Martínez-Jerónimo, F. Microcystis toxigenic strains in urban lakes: A case of study in Mexico City. Ecotoxicology 2010, 19, 1157–1165. [Google Scholar] [CrossRef]
- Alvarez-Hernandez, S.; De Lara-Isassi, G.; Arreguin-Espinoza, R.; Arreguin, B.; Hernandez-Santoyo, A.; Rodriguez-Romero, A. Isolation and partial characterization of giraffine, a lectin from the Mexican endemic alga Codium giraffa Silva. Bot. Mar. 1999, 42, 573–580. [Google Scholar] [CrossRef]
- Servín-Garcidueñas, L.E.; Martínez-Romero, E. Complete mitochondrial and plastid genomes of the green microalga Trebouxiophyceae sp. strain MX-AZ01 isolated from a highly acidic geothermal lake. Eukaryot. Cell 2012, 11, 1417–1418. [Google Scholar] [CrossRef]
- Pena-Pereira, F.; Kloskowski, A.; Namieśnik, J. Perspectives on the replacement of harmful organic solvents in analytical methodologies: A framework toward the implementation of a generation of eco-friendly alternatives. Green Chem. 2015, 17, 3687–3705. [Google Scholar] [CrossRef]
- Sosa-Hernández, J.; Escobedo-Avellaneda, Z.; Iqbal, H.; Welti-Chanes, J. State-of-the-Art Extraction Methodologies for Bioactive Compounds from Algal Biome to Meet Bio-Economy Challenges and Opportunities. Molecules 2018, 23, 2953. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.Y.; Lee, H.Y. Effect of Ultrasonic Extraction on Production and Structural Changes of C-Phycocyanin from Marine Spirulina maxima. Int. J. Mol. Sci. 2018, 19, 220. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Mazumder, A.; Giri, A.; Mishra, H.N. Optimization of Ultrasound-Assisted Extraction of β-Carotene from Chlorella Biomass (MCC7) and its Use in Fortification of Apple Jam. J. Food Process Eng. 2017, 40, e12321. [Google Scholar] [CrossRef]
- da Silva, M.F.; Casazza, A.A.; Ferrari, P.F.; Aliakbarian, B.; Converti, A.; Bezerra, R.P.; Porto, A.L.F.; Perego, P. Recovery of phenolic compounds of food concern from Arthrospira platensis by green extraction techniques. Algal Res. 2017, 25, 391–401. [Google Scholar] [CrossRef]
- Guldhe, A.; Singh, B.; Rawat, I.; Bux, F. Synthesis of biodiesel from Scenedesmus sp. by microwave and ultrasound assisted in situ transesterification using tungstated zirconia as a solid acid catalyst. Chem. Eng. Res. Des. 2014, 92, 1503–1511. [Google Scholar] [CrossRef]
- Lorenzen, J.; Igl, N.; Tippelt, M.; Stege, A.; Qoura, F.; Sohling, U.; Brück, T. Extraction of microalgae derived lipids with supercritical carbon dioxide in an industrial relevant pilot plant. Bioprocess Biosyst. Eng. 2017, 40, 911–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crampon, C.; Nikitine, C.; Zaier, M.; Lépine, O.; Tanzi, C.D.; Vian, M.A.; Chemat, F.; Badens, E. Oil extraction from enriched Spirulina platensis microalgae using supercritical carbon dioxide. J. Supercrit. Fluids 2017, 119, 289–296. [Google Scholar] [CrossRef]
- He, B.; Wang, Y.; Dou, X.; Chen, Y.F. Supercritical CO2 extraction of docosahexaenoic acid from Schizochytrium limacinum using vegetable oils as entrainer. Algal Res. 2017, 21, 58–63. [Google Scholar] [CrossRef]
- Albarelli, J.Q.; Santos, D.T.; Ensinas, A.V.; Marechal, F.; Cocero, M.J.; Meireles, M.A.A. Comparison of extraction techniques for product diversification in a supercritical water gasification-based sugarcane-wet microalgae biorefinery: Thermoeconomic and environmental analysis. J. Clean. Prod. 2018, 201, 697–705. [Google Scholar] [CrossRef]
- Araujo, G.S.; Matos, L.J.; Fernandes, J.O.; Cartaxo, S.J.; Gonçalves, L.R.; Fernandes, F.A.; Farias, W.R. Extraction of lipids from microalgae by ultrasound application: Prospection of the optimal extraction method. Ultrason. Sonochem. 2013, 20, 95–98. [Google Scholar] [CrossRef]
- Ma, Q.Y.; Fang, M.; Zheng, J.H.; Ren, D.F.; Lu, J. Optimised extraction of β-carotene from Spirulina platensis and hypoglycaemic effect in streptozotocin-induced diabetic mice. J. Sci. Food Agric. 2016, 96, 1783–1789. [Google Scholar] [CrossRef]
- Esquivel-Hernández, D.A.; Ibarra-Garza, I.P.; Rodríguez-Rodríguez, J.; Cuéllar-Bermúdez, S.P.; Rostro-Alanis, M.D.J.; Alemán-Nava, G.S.; García-Pérez, J.S.; Parra-Saldívar, R. Green extraction technologies for high-value metabolites from algae: A review. Biofuels Bioprod. Biorefining 2017, 11, 215–231. [Google Scholar] [CrossRef]
- Ma, Y.A.; Cheng, Y.M.; Huang, J.W.; Jen, J.F.; Huang, Y.S.; Yu, C.C. Effects of ultrasonic and microwave pretreatments on lipid extraction of microalgae. Bioprocess Biosyst. Eng. 2014, 37, 1543–1549. [Google Scholar] [CrossRef] [PubMed]
- Zinnai, A.; Sanmartin, C.; Taglieri, I.; Andrich, G.; Venturi, F. Supercritical fluid extraction from microalgae with high content of LC-PUFAs. A case of study: Sc-CO2 oil extraction from Schizochytrium sp. J. Supercrit. Fluids 2016, 116, 126–131. [Google Scholar] [CrossRef]
- Tavanandi, H.A.; Mittal, R.; Chandrasekhar, J.; Raghavarao, K.S.M.S. Simple and efficient method for extraction of C-Phycocyanin from dry biomass of Arthospira platensis. Algal Res. 2018, 31, 239–251. [Google Scholar] [CrossRef]
- Obeid, S.; Beaufils, N.; Camy, S.; Takache, H.; Ismail, A.; Pontalier, P.Y. Supercritical carbon dioxide extraction and fractionation of lipids from freeze-dried microalgae Nannochloropsis oculata and Chlorella vulgaris. Algal Res. 2018, 34, 49–56. [Google Scholar] [CrossRef]
- Kong, W.; Liu, N.; Zhang, J.; Yang, Q.; Hua, S.; Song, H.; Xia, C. Optimization of ultrasound-assisted extraction parameters of chlorophyll from Chlorella vulgaris residue after lipid separation using response surface methodology. J. Food Sci. Technol. 2014, 51, 2006–2013. [Google Scholar] [CrossRef]
- Ido, A.L.; de Luna, M.D.G.; Capareda, S.C.; Maglinao, A.L., Jr.; Nam, H. Application of central composite design in the optimization of lipid yield from Scenedesmus obliquus microalgae by ultrasonic-assisted solvent extraction. Energy 2018, 157, 949–956. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Allen, J.D.; Kanitkar, A.; Boldor, D. Oil extraction from Scenedesmus obliquus using a continuous microwave system–design, optimization, and quality characterization. Bioresour. Technol. 2011, 102, 3396–3403. [Google Scholar] [CrossRef]
- Pasquet, V.; Chérouvrier, J.R.; Farhat, F.; Thiéry, V.; Piot, J.M.; Bérard, J.B.; Kaas, R.; Serive, B.; Patrice, T.; Cadoret, J.P.; et al. Study on the microalgal pigments extraction process: Performance of microwave assisted extraction. Process Biochem. 2011, 46, 59–67. [Google Scholar] [CrossRef]
- Vinatoru, M.; Mason, T.J.; Calinescu, I. Ultrasonically assisted extraction (UAE) and microwave assisted extraction (MAE) of functional compounds from plant materials. TrAC Trends Anal. Chem. 2017, 97, 159–178. [Google Scholar] [CrossRef]
- Harris, J.; Viner, K.; Champagne, P.; Jessop, P.G. Advances in microalgal lipid extraction for biofuel production: A review. Biofuels Bioprod. Biorefining 2018, 12, 1118–1135. [Google Scholar] [CrossRef]
- Kumar, S.J.; Kumar, G.V.; Dash, A.; Scholz, P.; Banerjee, R. Sustainable green solvents and techniques for lipid extraction from microalgae: A review. Algal Res. 2017, 21, 138–147. [Google Scholar] [CrossRef]
- Mendes, R.L.; Nobre, B.P.; Cardoso, M.T.; Pereira, A.P.; Palavra, A.F. Supercritical carbon dioxide extraction of compounds with pharmaceutical importance from microalgae. Inorg. Chim. Acta 2003, 356, 328–334. [Google Scholar] [CrossRef]
- de Aguiar, A.C.; Osorio-Tobón, J.F.; Silva, L.P.S.; Barbero, G.F.; Martínez, J. Economic analysis of oleoresin production from malagueta peppers (Capsicum frutescens) by supercritical fluid extraction. J. Supercrit. Fluids 2018, 133, 86–93. [Google Scholar] [CrossRef]
- Hatami, T.; Johner, J.C.F.; Meireles, M.A.A. Extraction and fractionation of fennel using supercritical fluid extraction assisted by cold pressing. Ind. Crop. Prod. 2018, 123, 661–666. [Google Scholar] [CrossRef]
- Johner, J.C.; Hatami, T.; Meireles, M.A.A. Developing a supercritical fluid extraction method assisted by cold pressing for extraction of pequi (Caryocar brasiliense). J. Supercrit. Fluids 2018, 137, 34–39. [Google Scholar] [CrossRef]
- Solana, M.; Rizza, C.S.; Bertucco, A. Exploiting microalgae as a source of essential fatty acids by supercritical fluid extraction of lipids: Comparison between Scenedesmus obliquus, Chlorella protothecoides and Nannochloropsis salina. J. Supercrit. Fluids 2014, 92, 311–318. [Google Scholar] [CrossRef]
- Feller, R.; Matos, Â.P.; Mazzutti, S.; Moecke, E.H.; Tres, M.V.; Derner, R.B.; Oliveira, J.V.; Junior, A.F. Polyunsaturated ω-3 and ω-6 fatty acids, total carotenoids and antioxidant activity of three marine microalgae extracts obtained by supercritical CO 2 and subcritical n-butane. J. Supercrit. Fluids 2018, 133, 437–443. [Google Scholar] [CrossRef]
- Wei, F.; Gao, G.Z.; Wang, X.F.; Dong, X.Y.; Li, P.P.; Hua, W.; Wang, X.; Wu, X.M.; Chen, H. Quantitative determination of oil content in small quantity of oilseed rape by ultrasound-assisted extraction combined with gas chromatography. Ultrason. Sonochem. 2008, 15, 938–942. [Google Scholar] [CrossRef]
- Mercer, P.; Armenta, R.E. Developments in oil extraction from microalgae. Eur. J. Lipid Sci. Technol. 2011, 113, 539–547. [Google Scholar] [CrossRef]
- Klejdus, B.; Lojková, L.; Plaza, M.; Šnóblová, M.; Štěrbová, D. Hyphenated technique for the extraction and determination of isoflavones in algae: Ultrasound-assisted supercritical fluid extraction followed by fast chromatography with tandem mass spectrometry. J. Chromatogr. A 2010, 1217, 7956–7965. [Google Scholar] [CrossRef] [Green Version]
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Burgess, J.G.; Wright, P.C. Marine cyanobacteria—A prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar] [CrossRef]
- Shimizu, Y. Microalgal metabolites. Curr. Opin. Microbiol. 2003, 6, 236–243. [Google Scholar] [CrossRef]
- Reshef, V.; Mizrachi, E.; Maretzki, T.; Silberstein, C.; Loya, S.; Hizi, A.; Carmeli, S. New acylated sulfoglycolipids and digalactolipids and related known glycolipids from cyanobacteria with a potential to inhibit the reverse transcriptase of HIV-1. J. Nat. Prod. 1997, 60, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Hayashi, T.; Kojima, I. A natural sulfated polysaccharide, calcium spirulan, isolated from Spirulina platensis: In vitro and ex vivo evaluation of anti-herpes simplex virus and anti-human immunodeficiency virus activities. AIDS Res. Hum. Retrovir. 1996, 12, 1463–1471. [Google Scholar] [CrossRef]
- Moore, R.E.; Patterson, G.M.; Carmichael, W.W. New pharmaceuticals from cultured blue-green algae. Biomed. Importance Mar. Org. 1988, 13, 143–150. [Google Scholar]
- Dahms, H.-U.; Xu, Y.; Pfeiffer, C. Antifouling potential of cyanobacteria: A mini-review. Biofouling 2006, 22, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Piccaglia, R.; Marotti, M.; Grandi, S. Lutein and lutein ester content in different types of Tagetes patula and T. erecta. Ind. Crop. Prod. 1998, 8, 45–51. [Google Scholar] [CrossRef]
- Blanco, A.M.; Moreno, J.; Del Campo, J.A.; Rivas, J.; Guerrero, M.G. Outdoor cultivation of lutein-rich cells of Muriellopsis sp. in open ponds. Appl. Microbiol. Biotechnol. 2007, 73, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Choudhari, S.M.; Ananthanarayan, L.; Singhal, R.S. Use of metabolic stimulators and inhibitors for enhanced production of β-carotene and lycopene by Blakeslea trispora NRRL 2895 and 2896. Bioresour. Technol. 2008, 99, 3166–3173. [Google Scholar] [CrossRef]
- Sánchez, J.F.; Fernández-Sevilla, J.M.; Acién, F.G.; Cerón, M.C.; Pérez-Parra, J.; Molina-Grima, E. Biomass and lutein productivity of Scenedesmus almeriensis: Influence of irradiance, dilution rate and temperature. Appl. Microbiol. Biotechnol. 2008, 79, 719–729. [Google Scholar] [CrossRef]
- Fernández-Sevilla, J.M.; Fernández, F.A.; Grima, E.M. Biotechnological production of lutein and its applications. Appl. Microbiol. Biotechnol. 2010, 86, 27–40. [Google Scholar] [CrossRef]
- Cohen, Z. Production of polyunsaturated fatty acids (EPA, ARA, and GLA) by the microalgae Porphyridium and Spirulina. Ind. Appl. Single Cell Oils 1992, 243–273. [Google Scholar]
- Mao, T.K.; Water, J.V.D.; Gershwin, M.E. Effects of a Spirulina-based dietary supplement on cytokine production from allergic rhinitis patients. J. Med. Food 2005, 8, 27–30. [Google Scholar] [CrossRef]
- Barrow, C.; Shahidi, F. (Eds.) Marine Nutraceuticals and Functional Foods; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Becker, E.W. Microalgae: Biotechnology and Microbiology; Cambridge University Press: Cambridge, UK, 1994; Volume 10. [Google Scholar]
- Becker, W. Microalgae in Human and Animal Nutrition. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2004; Volume 312. [Google Scholar]
- Rasmussen, R.S.; Morrissey, M.T. Marine biotechnology for production of food ingredients. Adv. Food Nutr. Res. 2007, 52, 237–292. [Google Scholar]
- Garcı́a-González, A.; Ochoa, J.L. Anti-inflammatory activity of Debaryomyces hansenii Cu, Zn-SOD. Arch. Med. Res. 1999, 30, 69–73. [Google Scholar] [CrossRef]
- Guzmán-Murillo, M.A.; López-Bolaños, C.C.; Ledesma-Verdejo, T.; Roldan-Libenson, G.; Cadena-Roa, M.A.; Ascencio, F. Effects of fertilizer-based culture media on the production of exocellular polysaccharides and cellular superoxide dismutase by Phaeodactylum tricornutum (Bohlin). J. Appl. Phycol. 2007, 19, 33–41. [Google Scholar] [CrossRef]
- Priya, B.; Premanandh, J.; Dhanalakshmi, R.T.; Seethalakshmi, T.; Uma, L.; Prabaharan, D.; Subramanian, G. Comparative analysis of cyanobacterial superoxide dismutases to discriminate canonical forms. BMC Genom. 2007, 8, 435. [Google Scholar] [CrossRef] [PubMed]
- Thepenier, C.; Chaumont, D.; Gudin, C. Mass culture of Porphyridium cruentum: A multiproduct stategy for the biomass valorisation. In Algal Biotechnology; Stadler, T., Ed.; Elsevier Applied Science Publishers: London, UK, 1988; pp. 413–420. [Google Scholar]
- Antia, N.J.; Desai, I.D.; Romilly, M.J. The tocopherol, vitamin K, and related isoprenoid quinone composition of a unicellular red alga (Porphyridium cruentum). J. Phycol. 1970, 6, 305–312. [Google Scholar]
- Shi, X.M.; Chen, F. High yield production of lutein by heterotrophic Chlorella protothecoides in fed-batch systems. In Algae and Their Biotechnological Potential; Springer: Dordrecht, The Netherlands, 2001; pp. 107–119. [Google Scholar]
- Arya, V.; Gupta, V.K. A review on marine immunomodulators. Int. J. Pharm. Life Sci. 2011, 2, 751–758. [Google Scholar]
- Xiong, F.; Kopecky, J.; Nedbal, L. The occurrence of UV-B absorbing mycosporine-like amino acids in freshwater and terrestrial microalgae (Chlorophyta). Aquat. Bot. 1999, 63, 37–49. [Google Scholar] [CrossRef]
- Chu, W.L. Biotechnological applications of microalgae. IeJSME 2012, 6, S24–S37. [Google Scholar]
- Duval, B.; Shetty, K.; Thomas, W.H. Phenolic compounds and antioxidant properties in the snow alga Chlamydomonas nivalis after exposure to UV light. J. Appl. Phycol. 1999, 11, 559. [Google Scholar] [CrossRef]
- Karsten, U.; Lembcke, S.; Schumann, R. The effects of ultraviolet radiation on photosynthetic performance, growth and sunscreen compounds in aeroterrestrial biofilm algae isolated from building facades. Planta 2007, 225, 991–1000. [Google Scholar] [CrossRef]
- Matsukawa, R.; Hotta, M.; Masuda, Y.; Chihara, M.; Karube, I. Antioxidants from carbon dioxide fixing Chlorella sorokiniana. J. Appl. Phycol. 2000, 12, 263–267. [Google Scholar] [CrossRef]
- Barbosa, M.J.; Zijffers, J.W.; Nisworo, A.; Vaes, W.; van Schoonhoven, J.; Wijffels, R.H. Optimization of biomass, vitamins, and carotenoid yield on light energy in a flat-panel reactor using the A-stat technique. Biotechnol. Bioeng. 2005, 89, 233–242. [Google Scholar] [CrossRef]
- Egeland, E.S.; Guillard, R.R.; Liaaen-Jensen, S. Additional carotenoid prototype representatives and a general chemosystematic evaluation of carotenoids in Prasinophyceae (Chlorophyta). Phytochemistry 1997, 44, 1087–1097. [Google Scholar] [CrossRef]
- Cha, K.H.; Koo, S.Y.; Lee, D.U. Antiproliferative effects of carotenoids extracted from Chlorella ellipsoidea and Chlorella vulgaris on human colon cancer cells. J. Agric. Food Chem. 2008, 56, 10521–10526. [Google Scholar] [CrossRef]
- Leya, T.; Rahn, A.; Lütz, C.; Remias, D. Response of arctic snow and permafrost algae to high light and nitrogen stress by changes in pigment composition and applied aspects for biotechnology. FEMS Microbiol. Ecol. 2009, 67, 432–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramaraj, S.; Hemaiswarya, S.; Raja, R.; Ganesan, V.; Anbazhagan, C.; Carvalho, I.S.; Juntawong, N. Microalgae as an attractive source for biofuel production. In Environmental Sustainability; Springer: New Delhi, India, 2015; pp. 129–157. [Google Scholar]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Reitan, K.I.; Rainuzzo, J.R.; Olsen, Y. Effect of nutrient limitation on fatty acid and lipid content of marine microalgae 1. J. Phycol. 1994, 30, 972–979. [Google Scholar] [CrossRef]
- Cheng-Wu, Z.; Cohen, Z.; Khozin-Goldberg, I.; Richmond, A. Characterization of growth and arachidonic acid production of Parietochloris incisa comb. nov (Trebouxiophyceae, Chlorophyta). J. Appl. Phycol. 2002, 14, 453–460. [Google Scholar] [CrossRef]
- Ponomarenko, L.P.; Stonik, I.V.; Aizdaicher, N.A.; Orlova, T.Y.; Popovskaya, G.I.; Pomazkina, G.V.; Stonik, V.A. Sterols of marine microalgae Pyramimonas cf. cordata (Prasinophyta), Attheya ussurensis sp. nov.(Bacillariophyta) and a spring diatom bloom from Lake Baikal. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2004, 138, 65–70. [Google Scholar] [CrossRef]
- Cardozo, K.H.; Guaratini, T.; Barros, M.P.; Falcão, V.R.; Tonon, A.P.; Lopes, N.P.; Campos, S.; Torres, M.A.; Souza, A.O.; Colepicolo, P.; et al. Metabolites from algae with economical impact. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 146, 60–78. [Google Scholar] [CrossRef]
- Vilchez, C.; Garbayo, I.; Lobato, M.V.; Vega, J. Microalgae-mediated chemicals production and wastes removal. Enzym. Microb. Technol. 1997, 20, 562–572. [Google Scholar] [CrossRef]
- Uhlik, D.J.; Gowans, C.S. Synthesis of nicotinic acid in Chlamydomonas eugametos. Int. J. Biochem. 1974, 5, 79–84. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Commercial production of microalgae: Ponds, tanks, and fermenters. In Progress in Industrial Microbiology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 35, pp. 313–321. [Google Scholar]
- Running, J.A.; Severson, D.K.; Schneider, K.J. Extracellular production of L-ascorbic acid by Chlorella protothecoides, Prototheca species, and mutants of P. moriformis during aerobic culturing at low pH. J. Ind. Microbiol. Biotechnol. 2002, 29, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Carballo-Cárdenas, E.C.; Tuan, P.M.; Janssen, M.; Wijffels, R.H. Vitamin E (α-tocopherol) production by the marine microalgae Dunaliella tertiolecta and Tetraselmis suecica in batch cultivation. Biomol. Eng. 2003, 20, 139–147. [Google Scholar] [CrossRef]
- Chandra, R.; Parra, R.; MN Iqbal, H. Phycobiliproteins: A Novel Green Tool from Marine Origin Blue-Green Algae and Red Algae. Protein Pept. Lett. 2017, 24, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Du, S.W.; Zhang, L.K.; Han, K.; Chen, S.; Hu, Z.; Chen, W.; Hu, K.; Yin, L.; Wu, B.; Guan, Y.Q. Combined Phycocyanin and Hematoporphyrin Monomethyl Ether for Breast Cancer Treatment via Photosensitizers Modified Fe3O4 Nanoparticles Inhibiting the Proliferation and Migration of MCF-7 Cells. Biomacromolecules 2017, 19, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Koller, M.; Muhr, A.; Braunegg, G. Microalgae as versatile cellular factories for valued products. Algal Res. 2014, 6, 52–63. [Google Scholar] [CrossRef]
- Parmar, A.; Singh, N.K.; Kaushal, A.; Sonawala, S.; Madamwar, D. Purification, characterization and comparison of phycoerythrins from three different marine cyanobacterial cultures. Bioresour. Technol. 2011, 102, 1795–1802. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Wu, Y.; Wang, G.; Jia, T.; Zhang, Y. Purification and bioactivities of phycocyanin. Crit. Rev. Food Sci. Nutr. 2017, 57, 3840–3849. [Google Scholar] [CrossRef] [PubMed]
- Luengo, E.; Martínez, J.M.; Bordetas, A.; Álvarez, I.; Raso, J. Influence of the treatment medium temperature on lutein extraction assisted by pulsed electric fields from Chlorella vulgaris. Innov. Food Sci. Emerg. Technol. 2015, 29, 15–22. [Google Scholar] [CrossRef]
- Lin, J.H.; Lee, D.J.; Chang, J.S. Lutein production from biomass: Marigold flowers versus microalgae. Bioresour. Technol. 2015, 184, 421–428. [Google Scholar] [CrossRef]
- Del Campo, J.A.; García-González, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef]
- Gille, A.; Neumann, U.; Louis, S.; Bischoff, S.C.; Briviba, K. Microalgae as a potential source of carotenoids: Comparative results of an in vitro digestion method and a feeding experiment with C57BL/6J mice. J. Funct. Foods 2018, 49, 285–294. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B. Update on natural food pigments-A mini-review on carotenoids, anthocyanins, and betalains. Food Res. Int. 2018. [Google Scholar] [CrossRef]
- Chue, K.T.; Ten, L.N.; Oh, Y.K.; Woo, S.G.; Lee, M.; Yoo, S.A. Carotinoid compositions of five microalga species. Chem. Nat. Compd. 2012, 48, 141–142. [Google Scholar] [CrossRef]
- García, J.L.; de Vicente, M.; Galán, B. Microalgae, old sustainable food and fashion nutraceuticals. Microb. Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soni, R.A.; Sudhakar, K.; Rana, R.S. Spirulina–From growth to nutritional product: A review. Trends Food Sci. Technol. 2017, 69, 157–171. [Google Scholar] [CrossRef]
- Matos, J.; Cardoso, C.; Bandarra, N.M.; Afonso, C. Microalgae as healthy ingredients for functional food: A review. Food Funct. 2017, 8, 2672–2685. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, M.R.; Fernández, G.A.; Pérez, J.S.; Guerrero, J.G. Biomass nutrient profiles of the microalga Porphyridium cruentum. Food Chem. 2000, 70, 345–353. [Google Scholar] [CrossRef]
- Ovando, C.A.; Carvalho, J.C.D.; Vinícius de Melo Pereira, G.; Jacques, P.; Soccol, V.T.; Soccol, C.R. Functional properties and health benefits of bioactive peptides derived from Spirulina: A review. Food Rev. Int. 2018, 34, 34–51. [Google Scholar] [CrossRef]
- Batista, A.P.; Niccolai, A.; Fradinho, P.; Fragoso, S.; Bursic, I.; Rodolfi, L.; Biondi, N.; Tredici, M.R.; Sousa, I.; Raymundo, A. Microalgae biomass as an alternative ingredient in cookies: Sensory, physical and chemical properties, antioxidant activity and in vitro digestibility. Algal Res. 2017, 26, 161–171. [Google Scholar] [CrossRef]
- Ventura, S.P.M.; Nobre, B.P.; Ertekin, F.; Hayes, M.; Garciá-Vaquero, M.; Vieira, F.; Koc, M.; Gouveia, L.; Aires-Barros, M.R.; Palavra, A.M.F. Extraction of value-added compounds from microalgae. In Microalgae-Based Biofuels and Bioproducts; Elsevier: Woodhead Publishing, 2017; pp. 461–483. [Google Scholar]
- Markovits, A.; Conejeros, R.; López, L.; Lutz, M. Evaluation of marine microalga Nannochloropsis sp. as a potential dietary supplement. Chemical, nutritional and short term toxicological evaluation in rats. Nutr. Res. 1992, 12, 1273–1284. [Google Scholar]
- Rania, M.A.; Hala, M.T. Antibacterial and antifungal activity of Cyanobacteria and green Microalgae evaluation of medium components by Plackett-Burman design for antimicrobial activity of Spirulina platensis. Glob. J. Biotechnol. Biochem. 2008, 3, 22–31. [Google Scholar]
- Ruiz-Ruiz, F.; Mancera-Andrade, E.I.; Iqbal, H.M. Marine-derived bioactive peptides for biomedical sectors: A review. Protein Pept. Lett. 2017, 24, 109–117. [Google Scholar] [CrossRef] [PubMed]
- de Vera, C.; Díaz Crespín, G.; Hernández Daranas, A.; Montalvão Looga, S.; Lillsunde, K.E.; Tammela, P.; Perälä, M.; Hongisto, V.; Virtanen, J.; Rischer, H.; et al. Marine Microalgae: Promising Source for New Bioactive Compounds. Mar. Drugs 2018, 16, 317. [Google Scholar] [CrossRef]
- Bloor, S.; England, R.R. Elucidation and optimization of the medium constituents controlling antibiotic production by the cyanobacterium Nostoc muscorum. Enzym. Microb. Technol. 1991, 13, 76–81. [Google Scholar] [CrossRef]
- Galano, J.M.; Roy, J.; Durand, T.; Lee, J.C.Y.; Le Guennec, J.Y.; Oger, C.; Demion, M. Biological activities of non-enzymatic oxygenated metabolites of polyunsaturated fatty acids (NEO-PUFAs) derived from EPA and DHA: New anti-arrhythmic compounds? Mol. Asp. Med. 2018, 64, 161–168. [Google Scholar] [CrossRef]
- Abdulrazaq, M.; Innes, J.K.; Calder, P.C. Effect of ω-3 polyunsaturated fatty acids on arthritic pain: A systematic review. Nutrition 2017, 39, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Markworth, J.F.; Mitchell, C.J.; D’Souza, R.F.; Aasen, K.M.; Durainayagam, B.R.; Mitchell, S.M.; Chan, A.H.; Sinclair, A.J.; Garg, M.; Cameron-Smith, D. Arachidonic acid supplementation modulates blood and skeletal muscle lipid profile with no effect on basal inflammation in resistance exercise trained men. Prostaglandins Leukot. Essent. Fat. Acids 2018, 128, 74–86. [Google Scholar] [CrossRef]
- Habib, M.A.B. Review on Culture, Production and Use of Spirulina as Food for Humans and Feeds for Domestic Animals and Fish; Food and Agriculture Organization of the United Nations: Quebec, QC, Canada, 2008. [Google Scholar]
- Sonani, R.R.; Patel, S.; Bhastana, B.; Jakharia, K.; Chaubey, M.G.; Singh, N.K.; Madamwar, D. Purification and antioxidant activity of phycocyanin from Synechococcus sp. R42DM isolated from industrially polluted site. Bioresour. Technol. 2017, 245, 325–331. [Google Scholar] [CrossRef]
- Yan, G.A.; Jiang, J.W.; Wu, G.; Yan, X. Disappearance of linear alkylbenzene sulfonate from different cultures with Anabaena sp. HB 1017. Bull. Environ. Contam. Toxicol. 1998, 60, 329–334. [Google Scholar] [CrossRef]
- Radwan, S.S.; Al-Hasan, R.H. Oil pollution and cyanobacteria. In The Ecology of Cyanobacteria; Springer: The Netherlands, 2000; pp. 307–319. [Google Scholar]
- Raghukumar, C.; Vipparty, V.; David, J.; Chandramohan, D. Degradation of crude oil by marine cyanobacteria. Appl. Microbiol. Biotechnol. 2001, 57, 433–436. [Google Scholar]
- Radwan, S.S.; Al-Hasan, R.H.; Salamah, S.; Al-Dabbous, S. Bioremediation of oily sea water by bacteria immobilized in biofilms coating macroalgae. Int. Biodeterior. Biodegrad. 2002, 50, 55–59. [Google Scholar] [CrossRef]
- Mansy, A.E.R.; El-Bestawy, E. Toxicity and biodegradation of fluometuron by selected cyanobacterial species. World J. Microbiol. Biotechnol. 2002, 18, 125–131. [Google Scholar] [CrossRef]
- Uma, L.; Subramanian, G. Effective use of cyanobacteria in effluent treatment. In Proceedings of the National Symposium Cyanobacerial Nitrogen Fixation, IARI, New Delhi, India; 1990; pp. 437–444. [Google Scholar]
- Scherer, M.D.; de Oliveira, A.C.; Magalhães Filho, F.J.C.; Ugaya, C.M.L.; Mariano, A.B.; Vargas, J.V.C. Environmental study of producing microalgal biomass and bioremediation of cattle manure effluents by microalgae cultivation. Clean Technol. Environ. Policy 2017, 19, 1745–1759. [Google Scholar] [CrossRef]
- Matsunaga, T.; Sudo, H.; Takemasa, H.; Wachi, Y.; Nakamura, N. Sulfated extracellular polysaccharide production by the halophilic cyanobacterium Aphanocapsa halophytia immobilized on light-diffusing optical fibers. Appl. Microbiol. Biotechnol. 1996, 45, 24–27. [Google Scholar] [CrossRef]
- Dellamatrice, P.M.; Silva-Stenico, M.E.; de Moraes, L.A.B.; Fiore, M.F.; Monteiro, R.T.R. Degradation of textile dyes by cyanobacteria. Braz. J. Microbiol. 2017, 48, 25–31. [Google Scholar] [CrossRef]
- Pierre, G.; Zhao, J.M.; Orvain, F.; Dupuy, C.; Klein, G.L.; Graber, M.; Maugard, T. Seasonal dynamics of extracellular polymeric substances (EPS) in surface sediments of a diatom-dominated intertidal mudflat (Marennes–Oléron, France). J. Sea Res. 2014, 92, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Bhunia, B.; Uday, U.S.P.; Oinam, G.; Mondal, A.; Bandyopadhyay, T.K.; Tiwari, O.N. Characterization, genetic regulation and production of cyanobacterial exopolysaccharides and its applicability for heavy metal removal. Carbohydr. Polym. 2018, 179, 228–243. [Google Scholar] [CrossRef]
- Bilal, M.; Rasheed, T.; Sosa-Hernández, J.; Raza, A.; Nabeel, F.; Iqbal, H. Biosorption: An Interplay between Marine Algae and Potentially Toxic Elements—A Review. Mar. Drugs 2018, 16, 65. [Google Scholar] [CrossRef]
- Shuba, E.S.; Kifle, D. Microalgae to biofuels:‘Promising’alternative and renewable energy, review. Renew. Sustain. Energy Rev. 2018, 81, 743–755. [Google Scholar] [CrossRef]
- Khetkorn, W.; Rastogi, R.P.; Incharoensakdi, A.; Lindblad, P.; Madamwar, D.; Pandey, A.; Larroche, C. Microalgal hydrogen production–A review. Bioresour. Technol. 2017, 243, 1194–1206. [Google Scholar] [CrossRef]
- Batyrova, K.; Hallenbeck, P.C. Hydrogen production by a Chlamydomonas reinhardtii strain with inducible expression of photosystem II. Int. J. Mol. Sci. 2017, 18, 647. [Google Scholar] [CrossRef]
- Ainas, M.; Hasnaoui, S.; Bouarab, R.; Abdi, N.; Drouiche, N.; Mameri, N. Hydrogen production with the cyanobacterium Spirulina platensis. Int. J. Hydrog. Energy 2017, 42, 4902–4907. [Google Scholar] [CrossRef]
- Sengmee, D.; Cheirsilp, B.; Suksaroge, T.T.; Prasertsan, P. Biophotolysis-based hydrogen and lipid production by oleaginous microalgae using crude glycerol as exogenous carbon source. Int. J. Hydrog. Energy 2017, 42, 1970–1976. [Google Scholar] [CrossRef]
- Vargas, S.R.; dos Santos, P.V.; Zaiat, M.; do Carmo Calijuri, M. Optimization of biomass and hydrogen production by Anabaena sp. (UTEX 1448) in nitrogen-deprived cultures. Biomass Bioenergy 2018, 111, 70–76. [Google Scholar] [CrossRef]
- Nagarajan, D.; Lee, D.J.; Kondo, A.; Chang, J.S. Recent insights into biohydrogen production by microalgae–From biophotolysis to dark fermentation. Bioresour. Technol. 2017, 227, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Wünschiers, R.; Lindblad, P. Hydrogen in education—A biological approach. Int. J. Hydrog. Energy 2002, 27, 1131–1140. [Google Scholar] [CrossRef]
- Benemann, J.R. Hydrogen production by microalgae. J. Appl. Phycol. 2000, 12, 291–300. [Google Scholar] [CrossRef]
- Milano, J.; Ong, H.C.; Masjuki, H.H.; Chong, W.T.; Lam, M.K.; Loh, P.K.; Vellayan, V. Microalgae biofuels as an alternative to fossil fuel for power generation. Renew. Sustain. Energy Rev. 2016, 58, 180–197. [Google Scholar] [CrossRef]
- Park, J.Y.; Park, M.S.; Lee, Y.C.; Yang, J.W. Advances in direct transesterification of algal oils from wet biomass. Bioresour. Technol. 2015, 184, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Cho, J.M.; Chang, Y.K.; Oh, Y.K. Cell disruption and lipid extraction for microalgal biorefineries: A review. Bioresour. Technol. 2017, 244, 1317–1328. [Google Scholar] [CrossRef]
- Miao, X.; Wu, Q.; Yang, C. Fast pyrolysis of microalgae to produce renewable fuels. J. Anal. Appl. Pyrolysis 2004, 71, 855–863. [Google Scholar] [CrossRef]
- Willke, T.H.; Vorlop, K.D. Industrial bioconversion of renewable resources as an alternative to conventional chemistry. Appl. Microbiol. Biotechnol. 2004, 66, 131–142. [Google Scholar] [CrossRef]
- Gaurav, N.; Sivasankari, S.; Kiran, G.S.; Ninawe, A.; Selvin, J. Utilization of bioresources for sustainable biofuels: A Review. Renew. Sustain. Energy Rev. 2017, 73, 205–214. [Google Scholar] [CrossRef]
- Rizza, L.S.; Smachetti, M.E.S.; Do Nascimento, M.; Salerno, G.L.; Curatti, L. Bioprospecting for native microalgae as an alternative source of sugars for the production of bioethanol. Algal Res. 2017, 22, 140–147. [Google Scholar] [CrossRef]
- Harun, R.; Danquah, M.K.; Forde, G.M. Microalgal biomass as a fermentation feedstock for bioethanol production. J. Chem. Technol. Biotechnol. 2010, 85, 199–203. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Alberts, D.S.; Goldman, R.; Xu, M.J.; Dorr, R.T.; Quinn, J.; Welch, K.; Guillen-Rodriguez, J.; Aickin, M.; Peng, Y.M.; Loescher, L.; et al. Disposition and metabolism of topically administered α-tocopherol acetate: A common ingredient of commercially available sunscreens and cosmetics. Nutr. Cancer 1996, 26, 193–201. [Google Scholar] [CrossRef]
- Patel, H.M.; Rastogi, R.P.; Trivedi, U.; Madamwar, D. Structural characterization and antioxidant potential of phycocyanin from the cyanobacterium Geitlerinema sp. H8DM. Algal Res. 2018, 32, 372–383. [Google Scholar] [CrossRef]
- Camacho, F.G.; Rodríguez, J.G.; Mirón, A.S.; García, M.C.; Belarbi, E.H.; Chisti, Y.; Grima, E.M. Biotechnological significance of toxic marine dinoflagellates. Biotechnol. Adv. 2007, 25, 176–194. [Google Scholar] [CrossRef]
- Sathasivam, R.; Radhakrishnan, R.; Hashem, A.; Abd_Allah, E.F. Microalgae metabolites: A rich source for food and medicine. Saudi J. Biol. Sci. 2017. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Menakha, M. Pharmaceutical applications of cyanobacteria—A review. J. Acute Med. 2015, 5, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Nowruzi, B.; Haghighat, S.; Fahimi, H.; Mohammadi, E. Nostoc cyanobacteria species: A new and rich source of novel bioactive compounds with pharmaceutical potential. J. Pharm. Health Serv. Res. 2018, 9, 5–12. [Google Scholar] [CrossRef]
- Niedermeyer, T.H.J. Anti-infective natural products from Cyanobacteria. Planta Med. 2015, 81, 1309–1325. [Google Scholar] [CrossRef]
- Yaakob, Z.; Ali, E.; Zainal, A.; Mohamad, M.; Takriff, M.S. An overview: Biomolecules from microalgae for animal feed and aquaculture. J. Biol. Res.-Thessalon. 2014, 21, 6. [Google Scholar] [CrossRef]
- Sandoval, G.; Casas-Godoy, L.; Bonet-Ragel, K.; Rodrigues, J.; Ferreira-Dias, S.; Valero, F. Enzyme-Catalyzed Production of Biodiesel as Alternative to Chemical-Catalyzed Processes: Advantages and Constraints. Curr. Biochem. Eng. 2017, 4, 109–141. [Google Scholar] [CrossRef]
- Hosseini, N.S.; Shang, H.; Scott, J.A. Biosequestration of industrial off-gas CO2 for enhanced lipid productivity in open microalgae cultivation systems. Renew. Sustain. Energy Rev. 2018, 92, 458–469. [Google Scholar] [CrossRef]
- Packer, M. Algal capture of carbon dioxide; biomass generation as a tool for greenhouse gas mitigation with reference to New Zealand energy strategy and policy. Energy Policy 2009, 37, 3428–3437. [Google Scholar] [CrossRef]
- Kavitha, G.; Rengasamy, R.; Inbakandan, D. Polyhydroxybutyrate production from marine source and its application. Int. J. Biol. Macromol. 2018, 111, 102–108. [Google Scholar] [CrossRef]
- Archer, E.; Petrie, B.; Kasprzyk-Hordern, B.; Wolfaardt, G.M. The fate of pharmaceuticals and personal care products (PPCPs), endocrine disrupting contaminants (EDCs), metabolites and illicit drugs in a WWTW and environmental waters. Chemosphere 2017, 174, 437–446. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State | Municipality/Location | Microalgae | References |
|---|---|---|---|
| Baja California | Ensenada | Aphanocapsa marina | [19] |
| Komvophoron sp. | |||
| Phormidium sp. | |||
| Tetraselmis suecica | |||
| Heterococcus sp. | |||
| Amphora sp. (7) | |||
| Cymbella sp. (2) | |||
| Navicula sp. (4) | |||
| Diploneis sp. | |||
| Grammatophora angulosa | |||
| Synedra sp. | |||
| Veracruz | Catemaco | Aphanothece comasii | [17] |
| Cyanotetras aerotopa | |||
| Cylindrospermopsis catemaco | |||
| Cylindrospermopsis taverae | |||
| Planktolyngbya regularis | |||
| San Luis Potosí | Cyanobacterium lineatum | ||
| Puebla | Alchichica | Cyclotella alchichicana | |
| Chroococcus deltoids | |||
| Baja California, Colima, Michoacan, Guerrero, Tamaulipas, Veracruz, Hidalgo, Mexico city | Ensenada, Manzanillo, Lazaro Cardenas, Acapulco and Zihuatanejo, Laguna de Carpintero, Garrapatas and Barberena estuaries, Catemaco and Chalchoapan Lakes, Vicente Aguirre dam, Xochimilco Lake | Alexandrium tamarense Amphidinium sp. Cochlodinium polykrikoides Heterocapsa pigmea Gyrodinium instriatum Gymnodinium catenatum Karlodinium veneficum Prorocentrum gracile Prorocentrum micans Prorocentrum triestimum Prorocentrum mexicanum Prorocentrum rathymum Protoceratium reticulatum Scrippsiella trochoidea Bacillaria paxilifera Cylindrotheca closterium Pseudonitszchia delicatisima Chattonella marina | [21] |
| Mexico City | Mexico City | Spirulina maxima | [22] |
| Baja California Sur | La Paz | Rhabdonema sp. | [23] |
| Schizochytrium sp. | |||
| Nitzchia sp. | |||
| Navicula sp. | |||
| Grammatophora sp. | |||
| Mexico City | Mexico City | Spirulina platensis | [24] |
| Spirulina maxima | |||
| Queretaro | Not specified | Oscillatoria sp. | [25] |
| Guanajuato | Valle de Santiago | Actinastrum sp. | [26] |
| Baja California Sur | La Paz | Lyngbya sp. | [27] |
| Oscillatoria sp. | |||
| Microcoleus sp. | |||
| Anabaena sp. | |||
| Nuevo León | Apodaca | Scenedesmus sp. | [28] |
| Cadereyta | Chlorella sorokiniana | ||
| Campeche | El Carmen | Anabaena sp. | [29] |
| Oscillatoria sp. | |||
| Anabaena sp. | |||
| Cylindrospermopsis cuspis | |||
| Oaxaca | Zipolite | Dermocarpella sp. | [30] |
| Morelos | Tlaquiltenango | Nostoc sp. | [31] |
| Mexico City | Mexico City | Desmodesmus sp. | [32] |
| Coahuila | Cuatrociénegas | Scenedesmus sp. | [33] |
| Mexico City | Mexico City | Microcystis | [34] |
| Michoacan | Michoacan | Codium giraffa | [35] |
| Guerrero | Papanoa | Codium giraffa | [36] |
| Michoacán | Los Azufres | Trebouxiophyceae sp. | [37] |
| Compound(s) of Interest | Species | Extraction Technique | References |
|---|---|---|---|
| C-phycocyanin Pigments | Spirulina maxima | Ultrasound | [39] |
| β-carotene | Chlorella sp. | Ultrasound | [40] |
| Polyphenols Flavonoids | Spirulina platensis | Microwave and Ultrasound | [41] |
| Lipids | Scenedesmus sp. | Microwave | [42] |
| Lipids | Scenedesmus obliquus & Scenedesmus obtusiusculus | Supercritical-CO2 | [43] |
| Oil | Spirulina platensis | Supercritical-CO2 | [44] |
| Docosahexaenoic acid | Schizochytrium limacinum | Supercritical-CO2 -vegetable oil | [45] |
| Lipids, Carotenoids | Chlorella vulgaris | Supercritical-CO2 | [46] |
| Lipids | Chlorella vulgaris | Ultrasound & Bligh and Dyer method | [47] |
| β-carotene | Spirulina platensis | Ultrasound | [48] |
| Vitamins Phycocyanin Fatty Acids | Spirulina platensis | Microwave | [49] |
| Lipids | Chlorella sp. | Microwave and Ultrasound | [50] |
| Long-chain PUFAs | Schizochytrium sp. | Supercritical-CO2 | [51] |
| Carotenoids Fatty Acids | Spirulina platensis | Microwave and Supercritical-CO2 -etOH | [49] |
| C-phycocyanin | Spirulina platensis | Ultrasound | [52] |
| Neutral Lipids | Chlorella vulgaris & Nannochloropsis oculata | Supercritical-CO2 | [53] |
| Chlorophyll | Chlorella vulgaris | Ultrasound | [54] |
| Lipids | Scenedesmus obliquus | Ultrasound + solvent | [55] |
| Microalgae | Bioactive Compounds | Biological Activity | References |
|---|---|---|---|
| Oscillatoriaceae sp. | Malyngolide | Antibacterial | [70] |
| Lyngbyatoxins | PKC activator | ||
| Debromoaplysiatoxin | Inflammatory | ||
| Lyngbya majuscula | Curacin A | Microtubulin assembly inhibitors | [71] |
| Kalkitoxin | Sodium channel blocker | ||
| Cyclic polypeptide | Anti-HIV activity | ||
| Oscillatoria raoi | Acetylated sulfoglycolipids | Antiviral | [72] |
| Spirulina platensis | Spirulan | Antiviral | [73] |
| Nostocaceae sp. | Nostocyclamide | Antifungal | [74] |
| Chroococcaceae sp. | Kawaguchipeptin B | Antibacterial | [75] |
| Mycrosistis aeuregonosa | |||
| Scenedesmus sp. | Lutein | Anti-oxidant | [76,77,78,79,80] |
| Spirulina (Arthrospira) | γ-Linolenic acid (GLA) | The integrity of tissues, delay of aging | [81] |
| Spirulina (Arthrospira) | Phycocyanin | Antioxidant, anti-inflammatory | [15,82] |
| Tetracelmis suecica | α- tocopherol | Antioxidant | [15] |
| Chlorella sp. | Galactose, rhamnose, mannose, arabinose, N-acetyl glucosamide and N-acetyl galactosamine | Immune stimulatory activity | [83] |
| Spirulina platensis and Anabaena sp. | Proteins | [84,85,86] | |
| Anabaena sp. | Superoxide dismutase (SOD) | Antioxidant, anti-inflammatory | [87,88,89,90] |
| Spirulina sp. | Vitamin C; vitamin K; vitamins , A and E; α-tocopherol | Antioxidant; blood cell formation; blood clotting mechanism | [15,91] |
| Chlorella sp. | Lutein, zeaxanthin, canthaxanthin | Antioxidant | [15,92] |
| Lyngbya majuscula | Microlin- A | Immunosuppressive | [93] |
| Chlorella sorokiniana and Scenedesmus spp. | Mycosporine-like amino acids (MAA) | UV-screening agent; sunscreen | [94,95,96,97] |
| Chlorella sp. | α-carotene Astaxanthin | Lower risk of premature death | [98] |
| C. sorokiniana | β-carotene | Food colorant; antioxidant property; cancer preventive properties; prevent night blindness; prevent liver fibrosis | [99,100] |
| Tretraselmis spp. | Zeaxanthin | Protect eye cells; antioxidant activity; neutralizing the free radicals | [101,102] |
| Nitzschia spp. | Triglycerides and hydrocarbons | Biofuels | [95,103,104] |
| Tetraselmis spp. and T. suecica | Arachidonic acid (AA) Eicosapentaenoic acid (EPA) | Nutritional supplements, aquaculture feeds | [105,106] |
| T. suecica | Sterols | Antidiabetic; anticancer; anti- inflammatory; anti-photoaging; anti-obesity; anti-inflammatory; antioxidant activities | [107,108] |
| Chlorella spp. and C. sorokiniana | Vitamin B Vitamin C | Decrease fatigue; reducing depression; protect against heart disease; protect the skin; anticancer activity Protect against cardiovascular disease; prenatal health problems; prevent from the eye disease; protect against skin wrinkling | [85,99,109,110,111,112] |
| C. sorokiniana and T. suecica | Vitamin E | Protect against toxic pollutants; Premenstrual syndrome protects against eye disorders; anti- Alzheimer’s disease; anti- diabetic properties | [85,98,111,113] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sosa-Hernández, J.E.; Romero-Castillo, K.D.; Parra-Arroyo, L.; Aguilar-Aguila-Isaías, M.A.; García-Reyes, I.E.; Ahmed, I.; Parra-Saldivar, R.; Bilal, M.; Iqbal, H.M.N. Mexican Microalgae Biodiversity and State-Of-The-Art Extraction Strategies to Meet Sustainable Circular Economy Challenges: High-Value Compounds and Their Applied Perspectives. Mar. Drugs 2019, 17, 174. https://doi.org/10.3390/md17030174
Sosa-Hernández JE, Romero-Castillo KD, Parra-Arroyo L, Aguilar-Aguila-Isaías MA, García-Reyes IE, Ahmed I, Parra-Saldivar R, Bilal M, Iqbal HMN. Mexican Microalgae Biodiversity and State-Of-The-Art Extraction Strategies to Meet Sustainable Circular Economy Challenges: High-Value Compounds and Their Applied Perspectives. Marine Drugs. 2019; 17(3):174. https://doi.org/10.3390/md17030174
Chicago/Turabian StyleSosa-Hernández, Juan Eduardo, Kenya D. Romero-Castillo, Lizeth Parra-Arroyo, Mauricio A. Aguilar-Aguila-Isaías, Isaac E. García-Reyes, Ishtiaq Ahmed, Roberto Parra-Saldivar, Muhammad Bilal, and Hafiz M. N. Iqbal. 2019. "Mexican Microalgae Biodiversity and State-Of-The-Art Extraction Strategies to Meet Sustainable Circular Economy Challenges: High-Value Compounds and Their Applied Perspectives" Marine Drugs 17, no. 3: 174. https://doi.org/10.3390/md17030174