The Biological Deep Sea Hydrothermal Vent as a Model to Study Carbon Dioxide Capturing Enzymes

{kind=link}
Abstract
:1. Introduction
2. Carbon Dioxide in the Environments of Marine Hydrothermal Vents
3. Fixation and Assimilation of Carbon
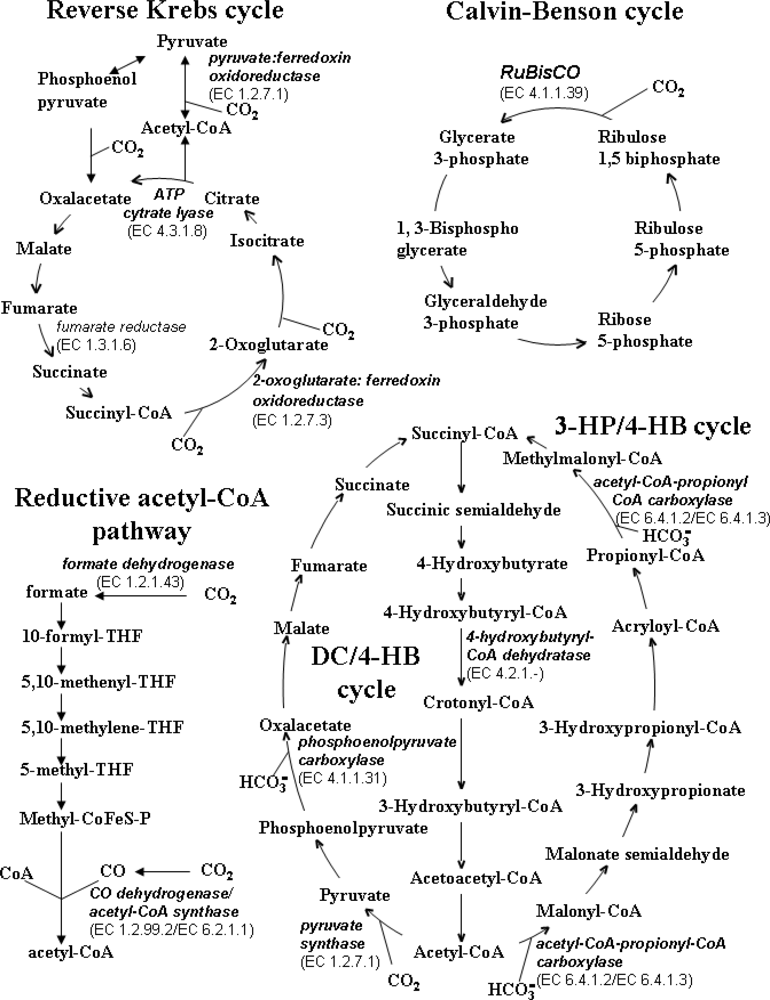
3.1. Calvin-Benson Cycle
3.2. Krebs Reverse Cycle (Reductive Tricarboxylic Acid Cycle)
3.3. 3-Hydroxypropionate Bicycle
3.4. Reductive Acetyl-CoA Pathway
3.5. Dicarboxylate/4-Hydroxybutyrate Cycle
3.6. 3-Hydroxypropionate/4-Hydroxybutyrate Cycle
3.7. Carbonic Anhydrase
4. Biotechnological Application
5. Conclusions
Acknowledgments
References
- Field, CB; Behrenfeld, MJ; Randerson, JT; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 28, 237–240. [Google Scholar]
- Falkowski, P; Scholes, RJ; Boyle, E; Canadell, J; Canfield, D; Elser, J; Gruber, N; Hibbard, K; Högberg, P; Linder, S; et al. The global carbon cycle: A test of our knowledge of earth as a system. Science 2000, 290, 291–296. [Google Scholar]
- Dubilier, N; Bergin, C; Lott, C. Symbiotic diversity in marine animals: The art of harnessing chemosynthesis. Nat Rev Microbiol 2008, 10, 725–740. [Google Scholar]
- Lonsdale, P. Clustering of suspension-feeding macrobenthos near abyssal hydrothermal vents at oceanic spreading centers. Deep Sea Res 1977, 24, 857–863. [Google Scholar]
- Corliss, JB; Dymond, J; Gordon, LI; Edmond, JM; Herzen, RPV; Ballard, RD; Green, K; Williams, D; Bainbridge, A; Crane, K; et al. Submarine thermal spring on the Galapagos Rift. Science 1979, 203, 1073–1083. [Google Scholar]
- Tunnicliffe, V; McArthur, A; McHugh, D. A biogeographical perspective of the deep-sea hydrothermal vent fauna. Adv Mar Biol 1998, 34, 353–442. [Google Scholar]
- Zierenberg, RA; Adams, MW; Arp, AJ. Life in extreme environments: Hydrothermal vents. Proc Natl Acad Sci USA 2000, 97, 12961–12962. [Google Scholar]
- Bott, MHP; Dean, DS. Stress diffusion from plate boundaries. Nature 1973, 243, 339–341. [Google Scholar]
- MacDonald, IR; Boland, GS; Baker, JS; Brooks, JM; Kennicutt, MC, II; Bidigare, RR. Gulf of Mexico hydrocarbon seep communities. II. Spatial distribution of seep organisms and hydrocarbons at Bush Hill. Mar Biol 1989, 101, 235–247. [Google Scholar]
- MacDonald, IR; Guinasso, NL; Reilly, JF; Brooks, JM; Callender, WR; Gabrielle, SG. Gulf of Mexico hydrocarbon seep communities. IV. Patterns in community structure and habitat. Geo-Mar Lett 1990, 10, 244–252. [Google Scholar]
- Felbeck, H. Chemoautotrophic potential of the hydrothermal vent tube worm Riftia pachyptila Jones (Vestimentifera). Science 1981, 213, 336–338. [Google Scholar]
- Felbeck, H; Childress, JJ; Somero, GN. Calvin-Benson cycle and sulfide oxidation enzymes in animals from sulfide rich environment habitats. Nature 1981, 293, 291–293. [Google Scholar]
- Cavanaugh, CM; Gardiner, SL; Jones, ML; Jannasch, HW; Waterbury, JB. Procaryotic cells in the hydrothermal vent tube worm Riftia pachyptila Jones: Possible chemoautotrophic symbionts. Science 1981, 213, 340–342. [Google Scholar]
- Gaill, F. Aspects of life development at deep sea hydrothermal vents. FASEB J 1993, 7, 558–565. [Google Scholar]
- Fisher, CR. Chemoautotrophic and methanotrophic symbioses in marine invertebrates. Aquat Sci 1996, 2, 399–436. [Google Scholar]
- Minic, Z; Hervé, G. Biochemical and enzymological aspects of the symbiosis between the deep-sea tubeworm Riftia pachyptila and its bacterial endosymbiont. Eur J Biochem 2004, 271, 3093–3102. [Google Scholar]
- Minic, Z; Serre, V; Hervé, G. Adaptation des organismes aux conditions extrêmes des sources hydrothermales marines profondes. Comp Rend Biol 2006, 329, 527–540. [Google Scholar]
- Desbruyères, D; Segonzac, M; Bright, M. Handbook of Deep-Sea Hydrothermal Vent Fauna, 2nd ed; Oberösterreichische Landesmuseen: Linz, Austria, 2006; pp. 1–544. [Google Scholar]
- Blöchl, E; Rachel, R; Burggraf, S; Hafenbradl, D; Jannasch, HW; Stetter, KO. Pyrolobus fumarii, gen. and sp. nov., represents a novel group of archaea, extending the upper temperature limit for life to 113 degrees C. Extremophiles 1997, 1, 14–21. [Google Scholar]
- Nakagawa, S; Takai, K. Deep-sea vent chemoautotrophs: Diversity, biochemistry and ecological significance. FEMS Microbiol Ecol 2008, 65, 1–14. [Google Scholar]
- Shively, JM; van Keulen, G; Meijer, WG. Something from almost nothing: Carbon dioxide fixation in chemoautotrophs. Annu Rev Microbiol 1998, 52, 191–230. [Google Scholar]
- Shively, JM; English, RS; Baker, SH; Cannon, GC. Carbon cycling: The prokaryotic contribution. Curr Opin Microbiol 2001, 3, 301–306. [Google Scholar]
- Berg, IA; Kockelkorn, D; Ramos-Vera, WH; Say, RF; Zarzycki, J; Hügler, M; Alber, BE; Fuchs, G. Autotrophic carbon fixation in archaea. Nat Rev Microbiol 2010, 6, 447–460. [Google Scholar]
- Hügler, M; Sievert, SM. Beyond the Calvin Cycle: Autotrophic carbon fixation in the ocean. Annu Rev Mar Sci 2011, 3, 261–289. [Google Scholar]
- Jannasch, HW; Wirsen, CO; Nelson, DC; Robertson, LA. Thiomicrospira crunogena sp. nov., a colorless, sulfur-oxidizing bacterium from a deep-sea hydrothermal vent. Int J Syst Bacteriol 1985, 35, 422–424. [Google Scholar]
- Goffredi, S; Childress, J; Desaulniers, N; Lee, R; Lallier, F; Hammond, D. Inorganic carbon acquisition by the hydrothermal vent tubeworm Riftia pachyptila depends upon high external P-CO2 and upon proton-equivalent ion transport by the worm. J Exp Biol 1997, 200, 883–896. [Google Scholar]
- Dobrinski, KP; Longo, DL; Scott, KM. The carbon-concentrating mechanism of the hydrothermal vent chemolithoautotroph Thiomicrospira crunogena. J Bacteriol 2005, 187, 5761–5766. [Google Scholar]
- Johnson, KS; Childress, JJ; Beehler, CL. Short term temperature variability in the Rose Garden hydrothermal vent field. Deep Sea Res 1988, 35, 1711–1722. [Google Scholar]
- Dobrinski, KP; Longo, DL; Scott, KM. The carbonconcentrating mechanism of the hydrothermal vent chemolithoautotroph Thiomicrospira crunogena. J Bacteriol 2005, 187, 5761–5766. [Google Scholar]
- Smith, KS; Ferry, JG. Prokaryotic carbonic anhydrases. FEMS Microbiol Rev 2000, 24, 335–366. [Google Scholar]
- De Cian, MC; Bailly, X; Morales, J; Strub, JM; Van Dorsselaer, A; Lallier, FH. Characterization of carbonic anhydrases from Riftia pachyptila, a symbiotic invertebrate from deep-sea hydrothermal vents. Proteins 2003, 51, 327–339. [Google Scholar]
- De Cian, MC; Andersen, AC; Bailly, X; Lallier, FH. Expression and localization of carbonic anhydrase and ATPases in the symbiotic tubeworm Riftia pachyptila. J Exp Biol 2003, 206, 399–409. [Google Scholar]
- Dobrinski, KP; Boller, AJ; Scott, KM. Expression and function of four carbonic anhydrase homologs in the deep-sea chemolithoautotroph Thiomicrospira crunogena. Appl Environ Microbiol 2010, 76, 3561–3567. [Google Scholar]
- Kochevar, RE; Govind, NS; Childress, JJ. Identification and characterization of two carbonic anhydrases from the hydrothermal vent tubeworm Riftia pachyptila Jones. Mol Mar Biol Biotechnol 1993, 2, 10–19. [Google Scholar]
- Minic, Z. Organisms of deep sea hydrothermal vents as a source for studying adaptation and evolution. Symbiosis 2009, 47, 121–132. [Google Scholar]
- Cavanaugh, C. Symbiotic chemoautotrophic bacteria in marine invertebrates from sulphide-rich habitats. Nature 1983, 302, 58–61. [Google Scholar]
- Karl, DM; Taylor, GT; Novitsky, JA; Jannasch, HW; Wirsen, CO; Pace, NR; Lane, DJ; Olsen, GJ; Giovannoni, SJ. A microbiological study of Guaymas Basin high temperature hydrothermal vents. Deep Sea Res 1988, 35, 777–791. [Google Scholar]
- Nelson, DC; Wirsen, CO; Jannasch, HW. Characterization of large, autotrophic Beggiatoa spp. abundant at hydrothermal vents of Guayamas basin. Appl Environ Microbiol 1989, 55, 2909–2917. [Google Scholar]
- Ruby, EG; Wirsen, CO; Jannasch, HW. Chemolithotrophic sulfur-oxidizing bacteria from the Galapagos rift hydrothermal vents. Appl Environ Microbiol 1981, 42, 317–324. [Google Scholar]
- Wirsen, CO; Jannasch, HW; Molyneaux, SJ. Chemosynthetic microbial activity at Mid-Atlantic Ridge hydrothermal vent sites. J Geophys Res 1993, 98, 9693–9703. [Google Scholar]
- Van Dover, CL; Fry, B. Stable isotopic compositions of hydrothermal vent organisms. Mar Biol 1989, 102, 257–263. [Google Scholar]
- Van Dover, CL; Humphris, SE; Fornari, D; Cavanaugh, CM; Collier, R; Goffredi, SK; Hashimoto, J; Lilley, MD; Reysenbach, AL; Shank, TM; et al. Biogeography and ecological setting of Indian Ocean hydrothermal vents. Science 2001, 294, 818–823. [Google Scholar]
- Elsaied, HE; Kimura, H; Naganuma, T. Composition of archaeal, bacterial, and eukaryal RuBisCO genotypes in three Western Pacific arc hydrothermal vent systems. Extremophiles 2007, 11, 191–202. [Google Scholar]
- Elsaied, H; Naganuma, T. Phylogenetic diversity of ribulose-1,5-bisphosphate carboxylase/oxygenase large-subunit genes from deep-sea microorganisms. Appl Environ Microbiol 2001, 67, 1751–1765. [Google Scholar]
- Eisen, JA; Nelson, KE; Paulsen, IT; Heidelberg, JF; Wu, M; Dodson, RJ; Deboy, R; Gwinn, ML; Nelson, WC; Haft, DH; et al. The complete genome sequence of Chlorobium tepidum TLS, a photosynthetic, anaerobic, green-sulfur bacterium. Proc Natl Acad Sci USA 2002, 99, 9509–9514. [Google Scholar]
- Finn, MW; Tabita, FR. Synthesis of catalytically active form III ribulose 1,5-bisphosphate carboxylase/oxygenase in archaea. J Bacteriol 2003, 185, 3049–3059. [Google Scholar]
- Badger, MR; Bek, EJ. Multiple Rubisco forms in proteobacteria: Their functional significance in relation to CO2 acquisition by the CBB cycle. J Exp Bot 2008, 59, 1525–1541. [Google Scholar]
- Akazawa, T; Takabe, T; Kobayashi, H. Molecular evolution of ribulose-l,5-bisphosphate carboxylase/oxgenase (RuBisCo). Trends Biochem Sci 1984, 9, 380–383. [Google Scholar]
- Newman, SM; Cattolico, RA. Ribulose bisphosphate earboxylase in evolution. Photosynth Res 1990, 26, 69–85. [Google Scholar]
- Tabita, FR. Microbial ribulose-1,5-bisphosphate carboxylase/oxygenase: A different perspective. Photosynth Res 1999, 60, 1–28. [Google Scholar]
- Giri, BJ; Bano, N; Hollibaugh, JT. Distribution of RuBisCO genotypes along a redox gradient in Mono Lake, California. Appl Environ Microbiol 2004, 70, 3443–3448. [Google Scholar]
- Kusian, B; Bowien, B. Organization and regulation of cbbCO2 assimilation genes in autotrophic bacteria. FEMS Microbiol Rev 1997, 21, 135–155. [Google Scholar]
- Tabita, FR; Hanson, TE; Li, H; Satagopan, S; Singh, J; Chan, S. Function, evolution, and structure of the Rubisco-like proteins and their Rubisco homologs. Microbiol Mol Biol Rev 2007, 71, 576–599. [Google Scholar]
- Tabita, FR; Satagopan, S; Hanson, TE; Kreel, NE; Scott, SS. Distinct form I, II, III, and IV Rubisco proteins from the three kingdoms of life provide clues about Rubisco evolution and structure/function relationships. J Exp Bot 2008, 59, 1515–1524. [Google Scholar]
- Watson, GM; Tabita, FR. Microbial ribulose 1,5-bisphosphate carboxylase/oxygenase: A molecule for phylogenetic and enzymological investigation. FEMS Microbiol Lett 1997, 146, 13–22. [Google Scholar]
- Elsaied, H; Sato, M; Naka, J; Naganuma, T. Analysis of 16S rRNA and RuBisCO large subunit genes from an abyssal low-temperature vent, Loihi Seamount, Hawaii. Cah Biol Mar 2002, 43, 403–408. [Google Scholar]
- Campbell, BJ; Cary, SC. Abundance of reverse tricarboxylic acid cycle genes in free-living microorganisms at deep sea hydrothermal vents. Appl Environ Microbiol 2004, 70, 6282–6289. [Google Scholar]
- Stein, JL; Haygood, M; Felbeck, H. Nucleotide sequence and expression of a deep-sea ribulose-1,5-bisphosphate carboxylase gene cloned from a chemoautotrophic bacterial endosymbiont. Proc Natl Acad Sci USA 1990, 87, 8850–8854. [Google Scholar]
- Kimura, H; Sato, M; Sasayama, Y; Naganuma, T. Molecular characterization and in situ localization of endosymbiotic 16S ribosomal RNA and RuBisCO genes in the Pogonophoran tissue. Mar Biotechnol 2003, 5, 261–269. [Google Scholar]
- Schwedock, J; Harmer, T; Scott, K; Hektor, H; Seitz, A; Fontana, M; Distel, D; Cavanaugh, C. Characterization and expression of genes from the RubisCO gene cluster of the chemoautotrophic symbiont of Solemya velum: cbbLSQO. Arch Microbiol 2004, 182, 18–29. [Google Scholar]
- Elsaied, H; Kimura, EH; Naganuma, T. Composition of archaeal, bacterial, and eukaryal RuBisCO genotypes in three Western Pacific arc hydrothermal vent systems. Extermofils 2006, 11, 191–202. [Google Scholar]
- Robinson, J; Stein, J; Cavanaugh, C. Cloning and sequencing of a form II ribulose-1,5-bisphosphate carboxylase/oxygenase from the bacterial symbiont of the hydrothermal vent tubeworm Riftia pachyptila. J Bacteriol 1998, 180, 1596–1599. [Google Scholar]
- Elsaied, H; Kimura, H; Naganuma, T. Molecular characterization and endosymbiotic localization of the gene encoding d-ribulose 1,5-bisphosphate carboxylase-oxygenase (RuBisCO) form II in the deep-sea vestimentiferan trophosome. Microbiology 2002, 148, 1947–1957. [Google Scholar]
- Kitano, K; Maeda, N; Fukui, T; Atomi, H; Imanaka, T; Miki, K. Crystal structure of a novel-type archaeal rubisco with pentagonal symmetry. Structure 2001, 9, 473–481. [Google Scholar]
- Klenk, HP; Clayton, RA; Tomb, JF; White, O; Nelson, KE; Ketchum, KA; Dodson, RJ; Gwinn, M; Hickey, EK; Peterson, JD; et al. The complete genome sequence of the hyperthermophilic, sulphate-reducing archaeon Archaeoglobus fulgidus. Nature 1997, 390, 364–370. [Google Scholar]
- Campbell, BJ; Stein, JL; Cary, SC. Evidence of chemolithoautotrophy in the bacterial community associated with Alvinella pompejana, a hydrothermal vent polychaete. Appl Environ Microbiol 2003, 69, 5070–5078. [Google Scholar]
- Campbell, BJ; Engel, AS; Porter, ML; Takai, K. The versatile epsilon-proteobacteria: Key players in sulphidic habitats. Nat Microbiol Rev 2006, 4, 458–468. [Google Scholar]
- Markert, S; Arndt, C; Felbeck, H; Becher, D; Sievert, SM; Hügler, M; Albrecht, D; Robidart, J; Bench, S; Feldman, RA; et al. Physiological proteomics of the uncultured endosymbiont of Riftia pachyptila. Science 2007, 315, 247–250. [Google Scholar]
- Ferrera, I; Longhorn, S; Banta, AB; Liu, Y; Preston, D; Reysenbach, AL. Diversity of 16S rRNA gene, ITS region and aclB gene of the Aquificales. Extremophiles 2007, 11, 57–64. [Google Scholar]
- Takaki, Y; Shimamura, S; Nakagawa, S; Fukuhara, Y; Horikawa, H; Ankai, A; Harada, T; Hosoyama, A; Oguchi, A; Fukui, S; et al. Bacterial lifestyle in a deep-sea hydrothermal vent chimney revealed by the genome sequence of the thermophilic bacterium Deferribacter desulfuricans SSM1. DNA Res 2010, 17, 123–137. [Google Scholar]
- Hügler, M; Wirsen, CO; Fuchs, G; Taylor, CD; Sievert, SM. Evidence for autotrophic CO2 fixation via the reductive tricarboxylic acid cycle by members of the epsilon subdivision of proteobacteria. J Bacteriol 2005, 187, 3020–3027. [Google Scholar]
- Hügler, M; Huber, H; Molyneaux, SJ; Vetriani, C; Sievert, SM. Autotrophic CO2 fixation via the reductive tricarboxylic acid cycle in different lineages within the phylum Aquificae: Evidence for two ways of citrate cleavage. Environ Microbiol 2007, 9, 81–92. [Google Scholar]
- Hügler, M; Gärtner, A; Imhoff, JF. Functional genes as markers for sulfur cycling and CO2 fixation in microbial communities of hydrothermal vents of the Logatchev field. FEMS Microbiol Ecol 2010, 73, 526–537. [Google Scholar]
- Takai, K; Campbell, BJ; Cary, SC; Suzuki, M; Oida, H; Nunoura, T; Hirayama, H; Nakagawa, S; Suzuki, Y; Inagaki, F; et al. Enzymatic and genetic characterization of carbon and energy metabolisms by deep-sea hydrothermal chemolithoautotrophic isolates of Epsilonproteobacteria. Appl Environ Microbiol 2005, 71, 7310–7320. [Google Scholar]
- Perner, M; Seifert, R; Weber, S; Koschinsky, A; Schmidt, K; Strauss, H; Peters, M; Haase, K; Imhoff, JF. Microbial CO2 fixation and sulfur cycling associated with low-temperature emissions at the Lilliput hydrothermal field, southern Mid-Atlantic Ridge (9 degrees S). Environ Microbiol 2007, 9, 1186–1201. [Google Scholar]
- Voordeckers, JW; Do, MH; Hügler, M; Ko, V; Sievert, SM; Vetriani, C. Culture dependent and independent analyses of 16S rRNA and ATP citrate lyase genes: A comparison of microbial communities from different black smoker chimneys on the Mid-Atlantic Ridge. Extremophiles 2008, 12, 627–640. [Google Scholar]
- Corre, E; Reysenbach, AL; Prieur, D. Epsilon proteobacterial diversity from a deep-sea hydrothermal vent on the Mid-Atlantic Ridge. FEMS Microbiol Lett 2001, 205, 329–335. [Google Scholar]
- Haddad, MA; Camacho, F; Durand, P; Cary, SC. Phlogenetic characterization of the epibiotic bacteria associated with the hydrothermal vent polychaete Alvinella pompejana. Appl Environ Microbiol 1995, 61, 1679–1687. [Google Scholar]
- Longnecker, K; Reysenbach, AL. Expansion of the geographic distribution of a novel lineage of epsilon Proteobacteria to a hydrothermal vent site on the Southern East Pacific Rise. FEMS Microbiol Ecol 2001, 35, 287–293. [Google Scholar]
- Moyer, CL; Dobbs, FC; Karl, DM. Phylogenetic diversity of the bacterial community from a microbial mat at an active, hydrothermal vent system, Loihi Seamount, Hawaii. Appl Environ Microbiol 1995, 61, 1555–1562. [Google Scholar]
- Reysenbach, AL; Longnecker, K; Kirshtein, J. Novel bacterial and archaeal lineages from an in situ growth chamber deployed at a Mid-Atlantic ridge hydrothermal vent. Appl Environ Microbiol 2000, 66, 3798–3806. [Google Scholar]
- Madigan, MT; Martinko, JM; Parker, J. Brock Biology of Microorganisms, 10th ed; Prentice Hall: Upper Saddle River, NJ, USA, 2002. [Google Scholar]
- Strauss, G; Fuchs, G. Enzymes of a novel autotrophic CO2-fixation pathway in the phototrophic bacterium Chloroflexus aurantiacus, the 3-hydroxypropionate cycle. Eur J Biochem 1993, 215, 633–643. [Google Scholar]
- Herter, S; Fuchs, G; Bacher, A; Eisenreich, W. A bicyclic autotrophic CO2 fixation pathway in Chloroflexus aurantiacus. J Biol Chem 2002, 277, 20277–20283. [Google Scholar]
- Zarzycki, J; Brecht, V; Muller, M; Fuchs, G. Identifying the missing steps of the autotrophic 3-hydroxypropionate CO2 fixation cycle in Chloroflexus aurantiacus. Proc Natl Acad Sci USA 2009, 106, 21317–21322. [Google Scholar]
- Pezacka, E; Wood, HG. Role of carbon monoxide dehydrogenase in the autotrophic pathway used by acetogenic bacteria. Proc Natl Acad Sci USA 1984, 81, 6261–6265. [Google Scholar]
- Ragsdale, SW; Wood, HG. Acetate biosynthesis by acetogenic bacteria. Evidence that carbon monoxide dehydrogenase is the condensing enzyme that catalyzes the final steps of the synthesis. J Biol Chem 1985, 260, 3970–3977. [Google Scholar]
- Ragsdale, SW; Pierce, E. Acetogenesis and the Wood-Ljungdahl pathway of CO2 fixation. Biochim Biophys Acta 2008, 1784, 1873–1898. [Google Scholar]
- Ljungdahl, LG. The autotrophic pathway of acetate synthesis in acetogenic bacteria. Annu Rev Microbiol 1986, 40, 415–450. [Google Scholar]
- Nakagawa, S; Takai, K. Methods for the isolation of thermophiles from deep-sea hydrothermal environments. Methods Microbiol 2006, 35, 55–91. [Google Scholar]
- Takai, K; Nakagawa, S; Reysenbach, AL; Hoek, J. Microbial ecology of Mid-Ocean Ridges and Back-arc Basins. In Back-Arc Spreading Systems: Geological, Biological, Chemical, and Physical Interactions; Christie, DM, Fisher, CR, Lee, SM, Givens, S, Eds.; American Geophysical Union: Washington DC, USA, 2006; pp. 185–213. [Google Scholar]
- Nakagawa, S; Inagaki, F; Suzuki, Y; Steinsbu, BO; Lever, MA; Takai, K; Engelen, B; Sako, Y; Wheat, CG; Horikoshi, K. Microbial community in black rust exposed to hot ridge flank crustal fluids. Appl Environ Microbiol 2006, 72, 6789–6799. [Google Scholar]
- Huber, H; Gallenberger, M; Jahn, U; Eylert, E; Berg, IA; Kockelkorn, D; Eisenreich, W; Fuchs, G. A dicarboxylate/4-hydroxybutyrate autotrophic carbon assimilation cycle in the hyperthermophilic Archaeum Ignicoccus hospitalis. Proc Natl Acad Sci USA 2008, 105, 7851–7856. [Google Scholar]
- Martins, BM; Dobbek, H; Cinkaya, I; Buckel, W; Messerschmidt, A. Crystal structure of 4-hydroxybutyryl-CoA dehydratase: Radical catalysis involving a [4Fe–4S] cluster and flavin. Proc Natl Acad Sci USA 2004, 101, 15645–15649. [Google Scholar]
- Buckel, W; Golding, GT. Radical enzymes in anaerobes. Annu Rev Microbiol 2006, 60, 27–49. [Google Scholar]
- Ramos-Vera, WH; Berg, IA; Fuchs, G. Autotrophic carbon dioxide assimilation in Thermoproteales revisited. J Bacteriol 2009, 191, 4286–4297. [Google Scholar]
- Paper, W; Jahn, U; Hohn, MJ; Kronner, M; Näther, DJ; Burghardt, T; Rachel, R; Stetter, KO; Huber, H. Ignicoccus hospitalis sp. nov., the host of “Nanoarchaeum equitans”. Int J Syst Evol Microbiol 2007, 57, 803–808. [Google Scholar]
- Zhou, H; Li, J; Peng, X; Meng, J; Wang, F; Ai, Y. Microbial diversity of a sulfide black smoker in main endeavour hydrothermal vent field, Juan de Fuca Ridge. J Microbiol 2009, 47, 235–247. [Google Scholar]
- Naganuma, T; Miyoshi, T; Kimura, H. Phylotype diversity of deep-sea hydrothermal vent prokaryotes trapped by 0.2- and 0.1-microm-pore-size filters. Extremophiles 2007, 11, 637–646. [Google Scholar]
- McCliment, EA; Voglesonger, KM; O’Day, PA; Dunn, EE; Holloway, JR; Cary, SC. Colonization of nascent, deep-sea hydrothermal vents by a novel Archaeal and Nanoarchaeal assemblage. Environ Microbiol 2006, 8, 114–125. [Google Scholar]
- Berg, IA; Kockelkorn, D; Buckel, W; Fuchs, G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 2007, 318, 1782–1786. [Google Scholar]
- Ishii, M; Miyake, T; Satoh, T; Sugiyama, H; Oshima, Y; Kodama, T; Igarashi, Y. Autotrophic carbon dioxide fixation in Acidianus brierleyi. Arch Microbiol 1997, 166, 368–371. [Google Scholar]
- Menendez, C; Bauer, Z; Huber, H; Gad’on, N; Stetter, KO; Fuchs, G. Presence of acetyl coenzyme A (CoA) carboxylase and propionyl-CoA carboxylase in autotrophic Crenarchaeota and indication for operation of a 3-hydroxypropionate cycle in autotrophic carbon fixation. J Bacteriol 1999, 181, 1088–1098. [Google Scholar]
- Hallam, SJ; Mincer, TJ; Schleper, C; Preston, CM; Roberts, K; Richardson, PM; DeLong, EF. Pathways of carbon assimilation and ammonia oxidation suggested by environmental genomic analyses of marine crenarchaeota. PLoS Biol 2006, 4, 520–536. [Google Scholar]
- Rusch, DB; Halpern, AL; Sutton, G. The Sorcerer II Global ocean sampling expedition: Northwest Atlantic through Eastern Tropical Pacific. PLoS Biol 2007, 5, e77. [Google Scholar]
- Yooseph, S; Sutton, G; Rusch, DB; Halpern, AL; Williamson, SJ; Remington, K; Eisen, JA; Heidelberg, KB; Manning, G; Li, W; et al. The sorcerer II global ocean sampling expedition: Expanding the universe of protein families. PLoS Biol 2007, 5, e16. [Google Scholar]
- Byrne, N; Strous, M; Crepeau, V; Kartal, B; Birrien, JL; Schmid, MC; Lesongeur, F; Schouten, S; Jaeschke, A; Jetten, M; et al. Presence and activity of anaerobic ammoniumoxidizing bacteria at deep-sea hydrothermal vents. ISME J 2009, 3, 117–123. [Google Scholar]
- Coleman, JR. The molecular and biochemical analyses of CO2 concentrating mechanism in cyanobacteria and microalgae. Plant Cell Environ 1991, 14, 861–867. [Google Scholar]
- Nimer, NA; Merrett, MJ. Calcification and utilization of inorganic carbon by the coccolithophorid Emiliania huxleyi Lohamnn. New Phytol 1992, 121, 173–177. [Google Scholar]
- Nimer, NA; Iglesias-Rodriguez, MD; Merrett, MJ. Bicarbonate utilization by marine phytoplankton species. J Phycol 1997, 33, 625–631. [Google Scholar]
- Badger, MR; Price, GD; Long, BM; Woodger, FJ. The environmental plasticity and ecological genomics of the cyanobacterial CO2 concentrating mechanism. J Exp Bot 2006, 57, 249–265. [Google Scholar]
- Price, GD; Badger, MR; Woodger, FJ; Long, BM. Advances in understanding the cyanobacterial CO2-concentrating-mechanism (CCM): Functional components, Ci transporters, diversity, genetic regulation and prospects for engineering into plants. J Exp Bot 2008, 59, 1441–1461. [Google Scholar]
- Dou, Z; Heinhorst, S; Williams, EB; Murin, CD; Shively, JM; Cannon, GC. CO2 fixation kinetics of Halothiobacillus neapolitanus mutant carboxysomes lacking carbonic anhydrase suggest the shell acts as a diffusional barrier for CO2. J Biol Chem 2008, 283, 10377–10384. [Google Scholar]
- Childress, JJ; Fischer, CR. The biology of hydrothermal vent animals: Physiology, biochemistry and autotrophic symbioses. Oceanogr Mar Biol Annu Rev 1992, 30, 337–341. [Google Scholar]
- Goffredi, SK; Childress, JJ; Lallier, FH; Desaulniers, NT. How to be the perfect host: CO2 and HS− accumulation and H+ elimination in the hydrothermal vent tube-worm Riftia pachyptila. Cah Biol Mar 1998, 39, 297–300. [Google Scholar]
- Scott, KM; Sievert, SM; Abril, FN; Ball, LA; Barrett, CJ; Blake, RA; Boller, AJ; Chain, PS; Clark, JA; Davis, CR; et al. The genome of deep-sea vent chemolithoautotroph Thiomicrospira crunogena. PLoS Biol 2006, 4, 1–17. [Google Scholar]
- Minic, Z; Simon, V; Penverne, B; Gaill, F; Herve, G. Contribution of the bacterial endosymbiont to the biosynthesis of pyrimidine nucleotides in the deep-sea tube worm Riftia pachyptila. J Biol Chem 2001, 276, 23777–23784. [Google Scholar]
- Minic, Z; Herve, G. Arginine metabolism in the deep sea tube worm Riftia pachyptila and its bacterial endosyrnbiont. J Biol Chem 2003, 278, 40527–40533. [Google Scholar]
- Debashish, G; Malay, S; Barindra, S; Joydeep, M. Marine enzymes. Adv Biochem Eng Biotechnol 2005, 96, 189–218. [Google Scholar]
- Guezennec, J. Les bactéries des sources hydrothermales profondes a l’origine de nouvelles molécules bioactives? In VertigO—La revue électronique en sciences de l'environnement; Les Éditions en environnement VertigO: Montréal, Canada, 2004; Volume 5, Numéro 3. [Google Scholar]
- Thornburg, CC; Zabriskie, TM; McPhail, KL. Deep-sea hydrothermal vents: Potential hot spots for natural products discovery? J Nat Prod 2010, 73, 489–499. [Google Scholar]
- Trincone, A. Potential biocatalysts originating from sea environments. J Mol Cat B: Enzym 2010, 66, 241–256. [Google Scholar]
- Lackne, KS. Climate Change: A guide to CO2 sequestration. Science 2003, 300, 1677–1678. [Google Scholar]
- Solomon, S; Plattner, GK; Knutti, R; Friedlingstein, P. Irreversible climate change due to carbon dioxide emissions. Proc Natl Acad Sci USA 2009, 106, 1704–1709. [Google Scholar]
- Schneider, SH; Thompson, S. Atmospheric CO2 and climate: The importance of the transient response. J Geophys Res 1981, 86, 3135–3147. [Google Scholar]
- Vitousek, PM; Mooney, HA; Lubchenco, J; Melillo, JM. Human Domination of Earth’s Ecosystems. Science 1997, 277, 494–499. [Google Scholar]
- Kumar, A; Ergas, S; Yuan, X; Sahu, A; Zhang, Q; Dewulf, J; Malcata, FX; van Langenhove, H. Enhanced CO2 fixation and biofuel production via microalgae: Recent developments and future directions. Trends Biotechnol 2010, 28, 371–380. [Google Scholar]
- Li, Y; Horsman, M; Wu, N; Lan, CQ; Dubios-Calero, M. Biofuels from microalgae. Biotechnol Prog 2008, 24, 815–820. [Google Scholar]
- Atsumi, S; Higashide, W; Liao, JC. Direct photosynthetic recycling of carbon dioxide to isobutyraldehyde. Nat Biotechnol 2009, 27, 1177–1180. [Google Scholar]
- Milne, JL; Cameron, JC; Page, LE; Benson, SM; Pakrasi, HB. Report from Workshop on Biological Capture and Utilization of CO2, Charles F Knight Center, Washington University in St. Louis, September 1–2, 2009; The Global Climate and Energy Project (GCEP): Stanford, CA, USA; pp. 1–43.
- Saini, R; Majhi, MC; Kapoor, R; Kumar, A; Kumar, R. CSD, a database of microbial strains for carbon fixation. Environ Modell Softw 2009, 24, 1133–1134. [Google Scholar]
- Wahlund, TM; Conway, T; Tabita, FR. Bioconversion of CO2 to ethanol and other compounds. Symposium on the Capture, Utilization and Disposal of CO2. Amer Chem Soc Div Fuel Chem 1996, 41, 1403–1406. [Google Scholar]
- Conway, T; Osman, YA; Konnan, JI; Hoffman, E; Ingram, LO. Promoter and nucleotide sequences of Zymomonas mobilis pyruvate decarboxylase. J Bacteriol 1987, 169, 949–954. [Google Scholar]
- Conway, T; Sewall, GW; Osman, YA; Ingram, LO. Cloning and sequencing of the alcohol dehydrogenase I1 gene from Zymomonas mobilis. J Bacteriol 1987, 169, 2591–2597. [Google Scholar]
- Ingram, LO; Conway, T. Expression of different levels of ethanologek enzymes from Zymomonas mobilis. J Bacteriol 1988, 169, 2591–2597. [Google Scholar]
- Dilmore, RM; Howard, BH; Soong, Y; Griffith, C; Hedges, SW; Degalbo, AD; Morreale, B; Baltrus, JP; Allen, DE; Fu, JK. Sequestration of CO2 in mixtures of caustic byproduct and saline waste water. Environ Eng Sci 2009, 26, 1325–1333. [Google Scholar]
- Mirjafari, P; Asghari, K; Mahinpey, N. Investigation the application of enzyme carbonic anhydrase for CO2 sequestration purposes. Ind Eng Chem Res 2007, 46, 921–926. [Google Scholar]
- Sharma, A; Bhattacharya, A; Pujari, R; Shrivastav, A. Characterization of carbonic anhydrase from diversified genus for biomimetic carbon-dioxide sequestration. Indian J Microbiol 2008, 48, 365–371. [Google Scholar]
- Bhattacharya, S; Nayak, A; Schiavone, M; Bhattacharya, SK. Solubilization and conversion of carbon dioxide: Novel spray reactors with immobilized carbonic anhydrase. Biotechnol Bioeng 2004, 86, 3–46. [Google Scholar]
- Borchert, M; Saunders, P. Heat-stable carbonic anhydrase and their use. WO Patent WO08/095057 2008. [Google Scholar]
- Lalande, JM; Tremblay, A. Process and a plant for the production of Portland cement clinker. US Patent 6,908,507 B2 2005. [Google Scholar]
- Lee, SW; Park, SB; Jeong, SK; Lim, KS; Lee, SH; Trachtenberg, MC. On carbon dioxide storage based on biomineralization strategies. Micron 2010, 41, 273–282. [Google Scholar]

© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Minic, Z.; Thongbam, P.D. The Biological Deep Sea Hydrothermal Vent as a Model to Study Carbon Dioxide Capturing Enzymes. Mar. Drugs 2011, 9, 719-738. https://doi.org/10.3390/md9050719
Minic Z, Thongbam PD. The Biological Deep Sea Hydrothermal Vent as a Model to Study Carbon Dioxide Capturing Enzymes. Marine Drugs. 2011; 9(5):719-738. https://doi.org/10.3390/md9050719
Chicago/Turabian StyleMinic, Zoran, and Premila D. Thongbam. 2011. "The Biological Deep Sea Hydrothermal Vent as a Model to Study Carbon Dioxide Capturing Enzymes" Marine Drugs 9, no. 5: 719-738. https://doi.org/10.3390/md9050719