Butyrolactone I Quantification from Lovastatin Producing Aspergillus terreus Using Tandem Mass Spectrometry—Evidence of Signalling Functions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Culture Conditions
2.3. Extraction of Butyrolactone I from the Cultures
2.4. Butyrolactone I Assay
2.4.1. HPLC
2.4.2. LC-ESI-MS-MS
2.5. Data Analysis
3. Results and Discussion
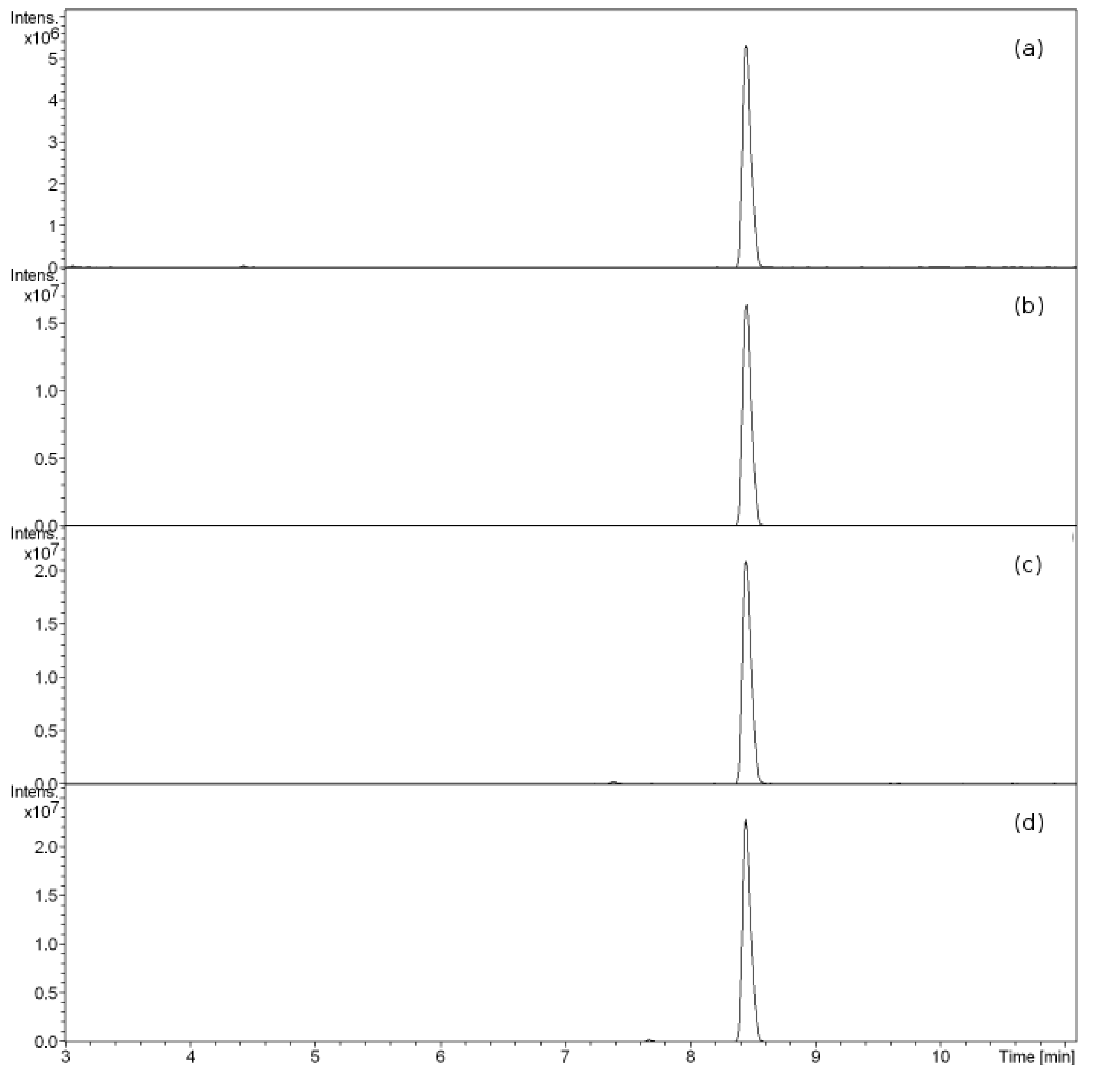
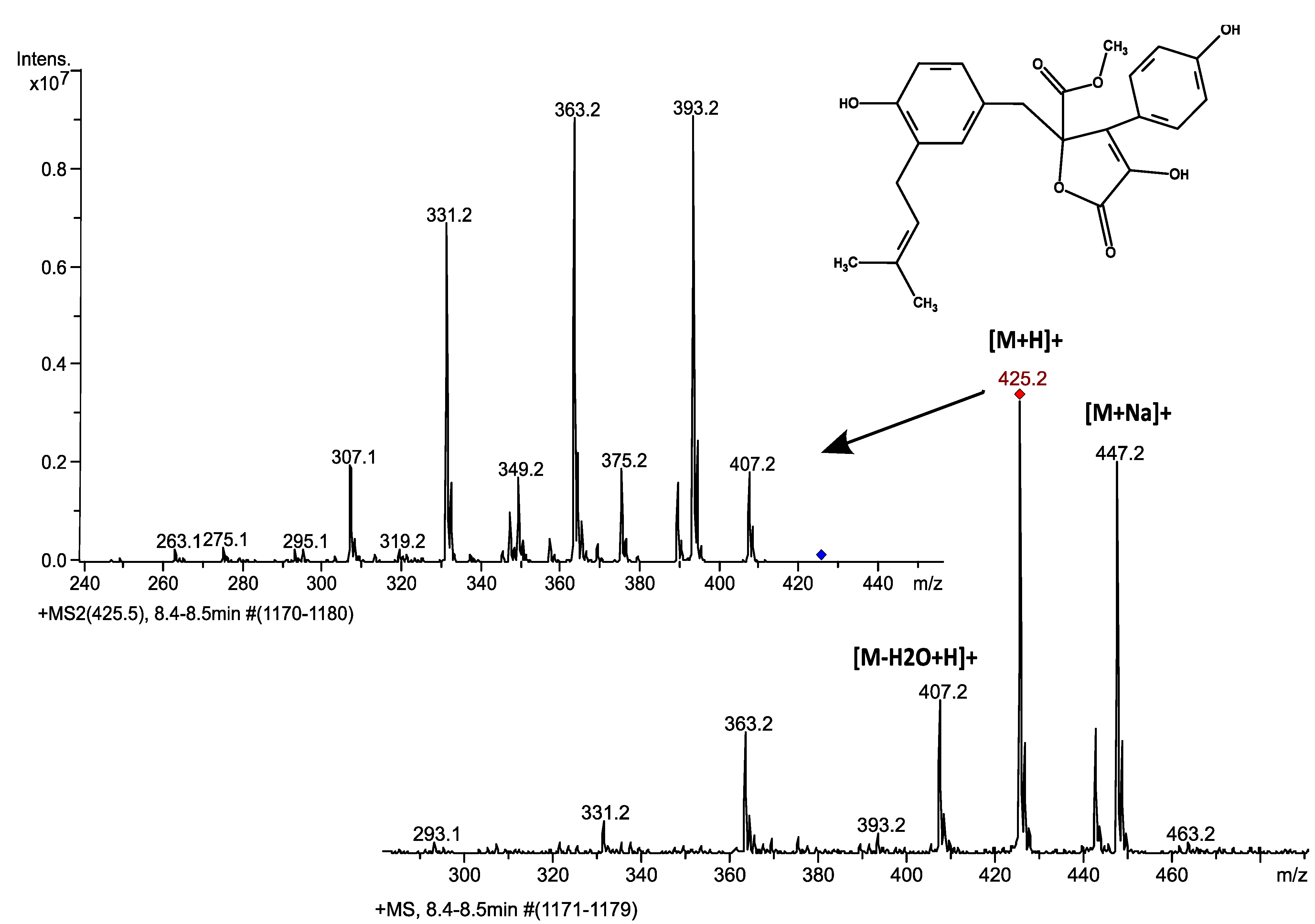
3.1. Butyrolactone I Fragmentation Pattern


3.2. Butyrolactone I Quantification


3.3. Supplementing with Butyrolactone I—Effects on Extracellular Concentration
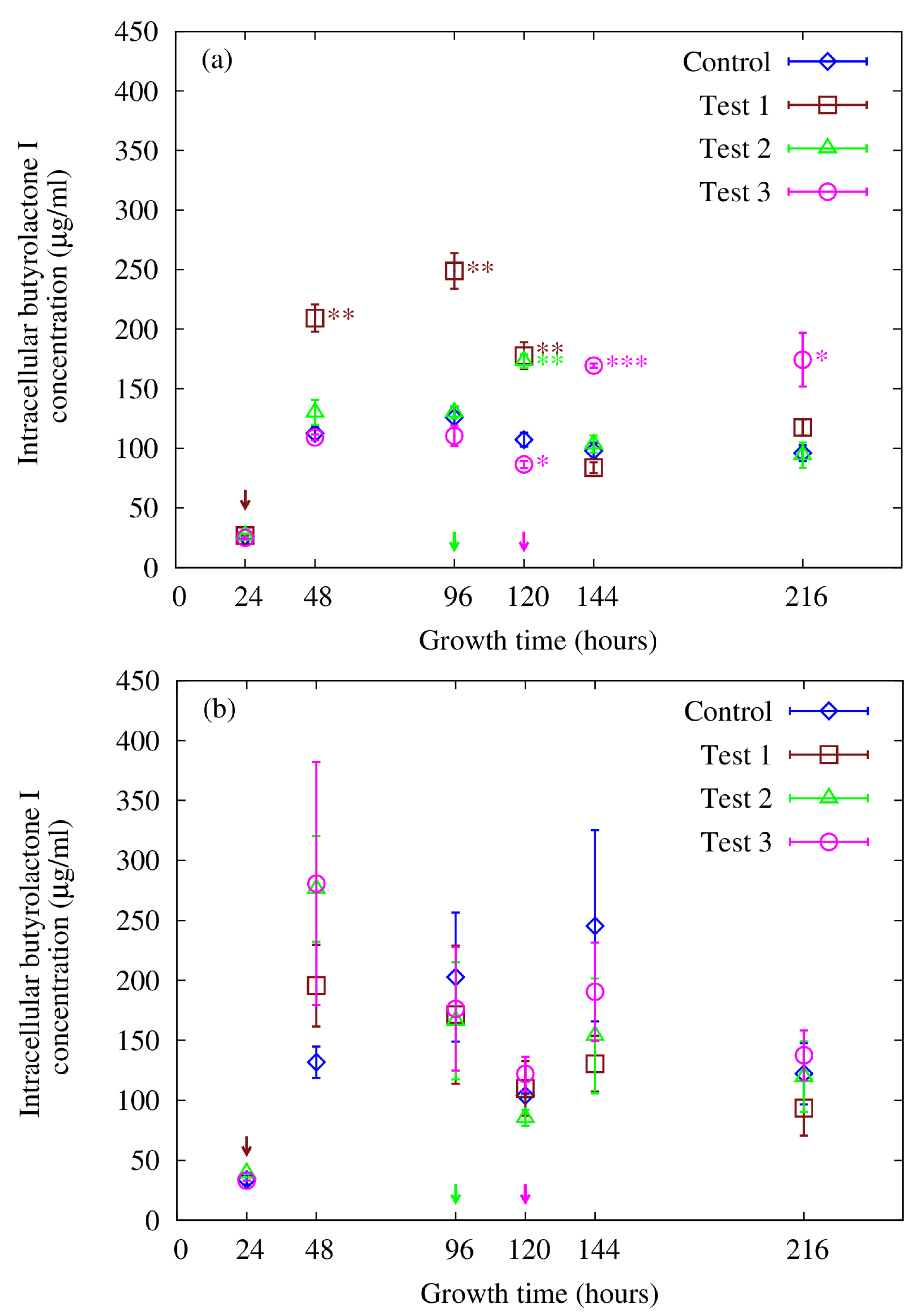
3.4. Supplementing with Butyrolactone I—Effects on Intracellular Concentration
3.5. Intracellular Accumulation of Butyrolactone I, Autoinduction and Quorum Sensing
3.6. Extracellular Butyrolactone I Is Depleted after 48 h Post Inoculation
3.7. Indications of Quorum Quenching?
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Alberts, A.W.; Chen, J.; Kuron, G.; Hunt, V.; Huff, J.; Hoffman, C.; Rothrock, J.; Lopez, M.; Joshua, H.; Harris, E.; et al. Mevinolin: A highly potent competitive inhibitor of hydroxymethylglutaryl-coenzyme A reductase and a cholesterol-lowering agent. Proc. Natl. Acad. Sci. USA 1980, 77, 3957–3961. [Google Scholar] [CrossRef]
- Arai, K.; Yoshimura, T.; Itatani, Y.; Yamamoto, Y. Metabolic products of Aspergillus terreus. VIII. Astepyrone: A novel metabolite of the strain IFO 4100. Chem. Pharm. Bull. 1983, 31, 925–933. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef]
- Kiriyama, N.; Nitta, K.; Sakaguchi, Y.; Taguchi, Y.; Yamamoto, Y. Studies on the metabolic products of Aspergillus terreus. III. Metabolites of the strain IFO 8835. (1). Chem. Pharm. Bull. 1977, 25, 2593–2601. [Google Scholar] [CrossRef]
- Lewis, R.E.; Wiederhold, N.P.; Lionakis, M.S.; Prince, R.A.; Kontoyiannis, D.P. Frequency and species distribution of gliotoxin-producing Aspergillus isolates recovered from patients at a tertiary-care cancer center. J. Clin. Microbiol. 2005, 43, 6120–6122. [Google Scholar] [CrossRef]
- Ojima, N.; Takenaka, S.; Seto, S. New butenolides from Aspergilllus terreus. Phytochemistry 1973, 12, 2527–2529. [Google Scholar] [CrossRef]
- Ojima, N.; Takenaka, S.; Seto, S. Structures of pulvinone derivatives from Aspergillus terreus. Phytochemistry 1975, 14, 573–576. [Google Scholar] [CrossRef]
- Ojima, N.; Takahashi, I.; Ogura, K.; Seto, S. New metabolites from Aspergillus terreus related to the biosynthesis of aspulvinones. Tetrahedron Lett. 1976, 17, 1013–1014. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Nishimura, K.-I.; Kiriyama, N. Studies on the metabolic products of Aspergillus terreus. I. Metabolites of the strain IFO 6123. Chem. Pharm. Bull. 1976, 24, 1853–1859. [Google Scholar]
- Okabe, M.; Lies, D.; Kanamasa, S.; Park, E.Y. Biotechnological production of itaconic acid and its biosynthesis in Aspergillus terreus. Appl. Micobiol. Biotechnol. 2009, 84, 597–606. [Google Scholar] [CrossRef]
- Lass-Flörl, C. The changing face of epidemiology of invasive fungal disease in Europe. Mycoses 2009, 52, 197–205. [Google Scholar] [CrossRef]
- Schimmel, T.G.; Coffman, A.D.; Parsons, S.J. Effect of butyrolactone I on the producing fungus, Aspergillus terreus. Appl. Environ. Microbiol. 1998, 64, 3707–3712. [Google Scholar]
- Askenazi, M.; Driggers, E.M.; Holtzman, D.A.; Norman, T.C.; Iverson, S.; Zimmer, D.P.; Boers, M.-E.; Blomquist, P.R.; Martinez, E.J.; Monreal, A.W.; et al. Integrating transcriptional and metabolite profiles to direct the engineering of lovastatin-producing fungal strains. Nat. Biotechnol. 2003, 21, 150–156. [Google Scholar] [CrossRef]
- Bizukojc, M.; Pawlowska, B.; Ledakowicz, S. Supplementation of the cultivation media with B-group vitamins enhances lovastatin biosynthesis by Aspergillus terreus. J. Biotechnol. 2007, 127, 258–268. [Google Scholar] [CrossRef]
- Bizukojc, M.; Ledakowicz, S. Simultaneous biosynthesis of (+)-geodin by a lovastatin-producing fungus Aspergillus terreus. J. Biotechnol. 2007, 132, 453–460. [Google Scholar] [CrossRef]
- Bizukojc, M.; Ledakowicz, S. Biosynthesis of lovastatin and (+)-geodin by Aspergillus terreus in batch and fed-batch culture in the stirred tank bioreactor. Biochem. Eng. J. 2008, 42, 198–207. [Google Scholar] [CrossRef]
- Raina, S.; de Vizio, D.; Palonen, E.K.; Odell, M.; Brandt, A.M.; Soini, J.T.; Keshavarz, T. Is quorum sensing involved in lovastatin production in the filamentous fungus Aspergillus terreus? Process Biochem. 2012, 47, 843–852. [Google Scholar] [CrossRef]
- Kitagawa, M.; Okabe, T.; Ogino, H.; Matsumoto, H.; Suzukitakahashi, I.; Kokubo, T.; Higashi, H.; Saitoh, S.; Taya, Y.; Yasuda, H.; et al. Butyrolactone I, a selective inhibitor of cdk2 and cdc2 kinase. Oncogene 1993, 8, 2425–2432. [Google Scholar]
- Nishio, K.; Ishida, T.; Arioka, H.; Kurokawa, H.; Fukuoka, K.; Nomoto, T.; Fukumoto, H.; Yokote, H.; Saijo, N. Antitumour effects of butyrolactone I, a selective cdc2 kinase inhibitor, on human lung cancer cell lines. Anticancer Res. 1996, 16, 3387–3395. [Google Scholar]
- Suzuki, M.; Hosaka, Y.; Matsushima, H.; Goto, T.; Kitamura, T.; Kawabe, K. Butyrolactone I induces cyclin B1 and causes G2/M arrest and skipping of mitosis in human prostate cell lines. Cancer Lett. 1999, 138, 121–130. [Google Scholar] [CrossRef]
- Wada, M.; Hosotani, R.; Lee, J.U.; Doi, R.; Koshiba, T.; Fujimoto, K.; Miyamoto, Y.; Tsuji, S.; Nakajima, S.; Okuyama, A.; et al. An exogenous cdk inhibitor, butyrolactone-I, induces apoptosis with increased Bax/Bcl-2 ratio in p53-mutated pancreatic cancer cells. Anticancer Res. 1998, 18, 2559–2566. [Google Scholar]
- Horinouchi, S. Mining and polishing of the treasure trove in the bacterial genus Streptomyces. Biosci. Biotechnol. Biochem. 2007, 71, 283–299. [Google Scholar] [CrossRef]
- Atkinson, S.; Williams, P. Quorum sensing and social networking in the microbial world. J. R. Soc. Interface 2009, 6, 959–978. [Google Scholar] [CrossRef]
- Hornby, J.M.; Jensen, E.C.; Lisec, A.D.; Tasto, J.J.; Jahnke, B.; Shoemaker, R.; Dussault, P.; Nickerson, K.W. Quorum sensing in the dimorphic fungus Candida albicans is mediated by farnesol. Appl. Environ. Microbiol. 2001, 67, 2982–2992. [Google Scholar] [CrossRef]
- Horowitz Brown, S.; Zarnowski, R.; Sharpee, W.C.; Keller, N.P. Morphological transitions governed by density dependence and lipoxygenase activity in Aspergillus flavus. Appl. Environ. Microbiol. 2008, 74, 5674–5685. [Google Scholar] [CrossRef]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef]
- Surette, M.G.; Bassler, B.L. Quorum sensing in Escherichia coli and Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 1998, 95, 7046–7050. [Google Scholar] [CrossRef]
- Schuster, M.; Lostroh, C.P.; Ogi, T.; Greenberg, E.P. Identification, timing, and signal specificity of Pseudomonas aeruginosa quorum-controlled genes: A transcriptome analysis. J. Bacteriol. 2003, 185, 2066–2079. [Google Scholar] [CrossRef]
- Raina, S.; de Vizio, D.; Odell, M.; Clements, M.; Vanhulle, S.; Keshavarz, T. Microbial quorum sensing: A tool or a target for antimicrobial therapy? Biotechnol. Appl. Biochem. 2009, 54, 65–84. [Google Scholar]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef]
- Schimmel, T.G.; Parsons, S.J. High purity, high yield procedure for butyrolactone I production from Aspergillus terreus. Biotechnol. Tech. 1999, 13, 379–384. [Google Scholar] [CrossRef]
- Sorrentino, F.; Roy, I.; Keshavarz, T. Impact of linoleic acid supplementation on lovastatin production in Aspergillus terreus cultures. Appl. Microbiol. Biotechnol. 2010, 88, 65–73. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; ISBN 3-900051-07-0. [Google Scholar]
- Ando, N.; Ueda, K.; Horinouchi, S. A Streptomyces griseus gene (sgaA) suppresses the growth disturbance caused by high osmolality and a high concentration of A-factor during early growth. Microbiology 1997, 143, 2715–2723. [Google Scholar] [CrossRef]
- Nitta, K.; Fujita, N.; Yoshimura, T.; Arai, K.; Yamamoto, Y. Metabolic products of Aspergillus terreus. IX. Biosynthesis of butyrolactone derivatives isolated from strains IFO 8835 and 4100. Chem. Pharm. Bull. 1983, 31, 1528–1533. [Google Scholar] [CrossRef]
- Uroz, S.; Dessaux, Y.; Oger, P. Quorum sensing and quorum quenching: The yin and yang of bacterial communication. ChemBioChem 2009, 10, 205–216. [Google Scholar] [CrossRef]
- Khan, S.R.; Farrand, S.K. The BlcC (AttM) lactonase of Agrobacterium tumefaciens does not quench the quorum-sensing system that regulates Ti plasmid conjugative transfer. J. Bacteriol. 2009, 191, 1320–1329. [Google Scholar] [CrossRef]
- Park, S.-Y.; Kang, H.-O.; Jang, H.-S.; Lee, J.-K.; Koo, B.-T.; Yum, D.-Y. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl. Environ. Microbiol. 2005, 71, 2632–2641. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Palonen, E.K.; Neffling, M.-R.; Raina, S.; Brandt, A.; Keshavarz, T.; Meriluoto, J.; Soini, J. Butyrolactone I Quantification from Lovastatin Producing Aspergillus terreus Using Tandem Mass Spectrometry—Evidence of Signalling Functions. Microorganisms 2014, 2, 111-127. https://doi.org/10.3390/microorganisms2020111
Palonen EK, Neffling M-R, Raina S, Brandt A, Keshavarz T, Meriluoto J, Soini J. Butyrolactone I Quantification from Lovastatin Producing Aspergillus terreus Using Tandem Mass Spectrometry—Evidence of Signalling Functions. Microorganisms. 2014; 2(2):111-127. https://doi.org/10.3390/microorganisms2020111
Chicago/Turabian StylePalonen, Elina K., Milla-Riina Neffling, Sheetal Raina, Annika Brandt, Tajalli Keshavarz, Jussi Meriluoto, and Juhani Soini. 2014. "Butyrolactone I Quantification from Lovastatin Producing Aspergillus terreus Using Tandem Mass Spectrometry—Evidence of Signalling Functions" Microorganisms 2, no. 2: 111-127. https://doi.org/10.3390/microorganisms2020111