
Toxicity of Plant Secondary Metabolites Modulating Detoxification Genes Expression for Natural Red Palm Weevil Pesticide Development



Abstract
:1. Introduction
2. Results
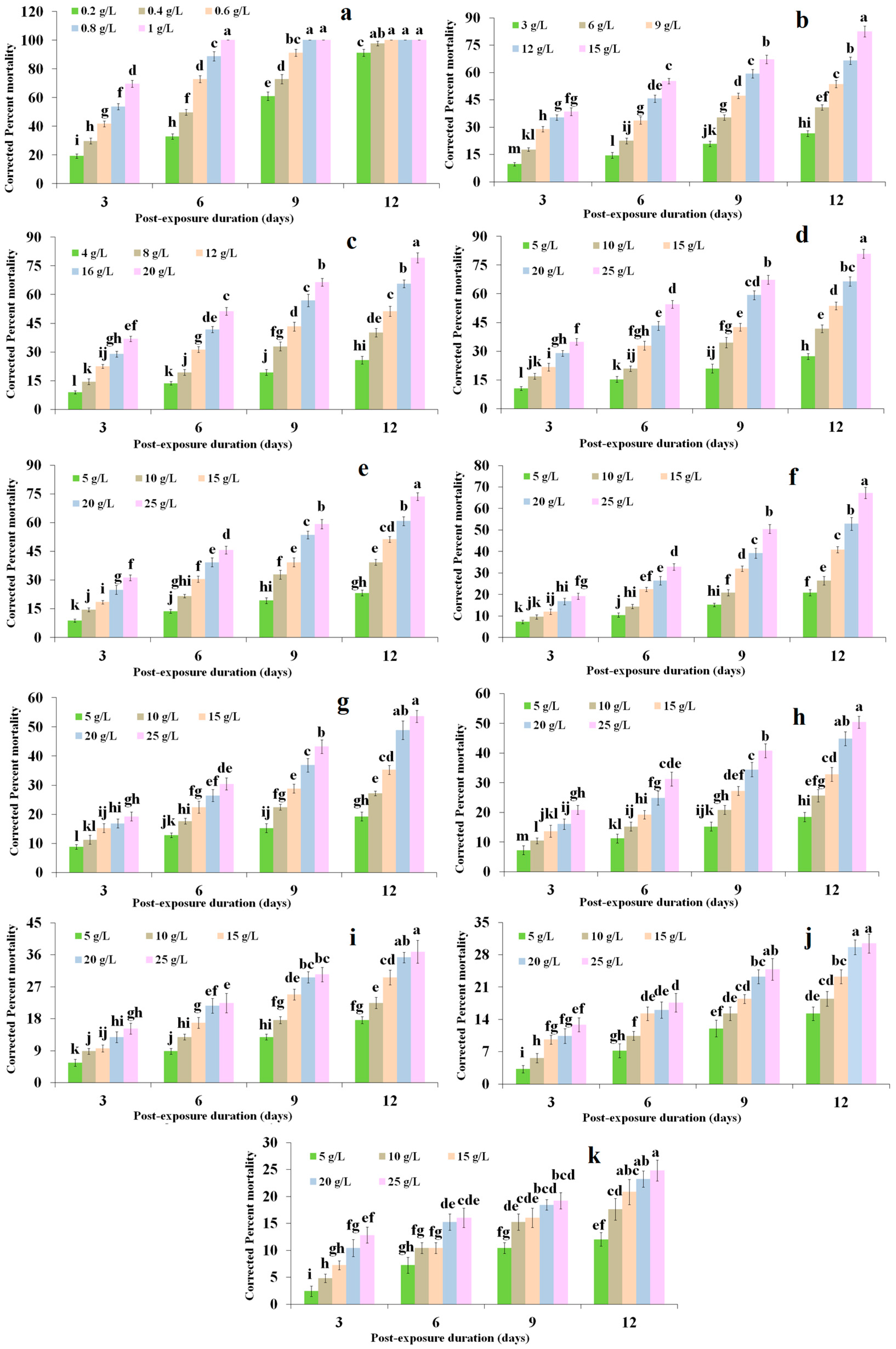
2.1. Dose Mortality Response
2.2. Growth Inhibition Activities of Red Palm Weevil Larvae
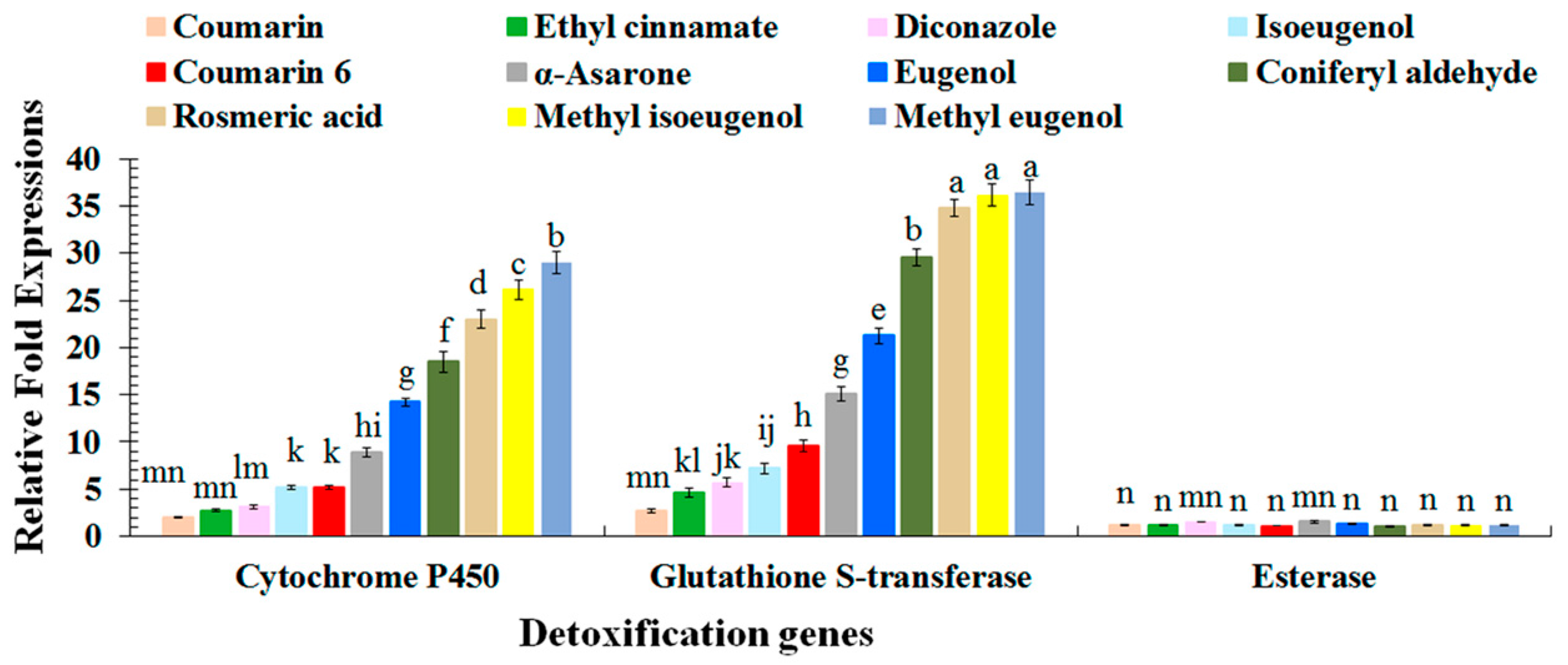
2.3. Quantitative Reverse Transcription Polymerase Chain Reaction Analyses of Red Palm Weevil Detoxification Genes
3. Discussion
4. Materials and Methods
4.1. Insects
4.2. Phenylpropanoid Plant Secondary Metabolites
4.3. Toxicity Bioassays
4.4. Nutritiopnal Indices of Red Palm Weevils
4.5. Host Detoxification Mechanism
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hussain, A.; Rizwan-ul-haq, M.; Al-jabr, A.M. Red palm weevil: Understanding the fungal disease mechanism and host defense. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Formatex Research Center: Badajoz, Spain, 2013; pp. 1278–1286. [Google Scholar]
- Hussain, A.; Rizwan-ul-Haq, M.; Al-Jabr, A.M.; Al-Ayied, H.Y. Managing invasive populations of red palm weevil: A worldwide perspective. J. Food Agric. Environ. 2013, 11, 456–463. [Google Scholar]
- Al-Ayedh, H.; Hussain, A.; Rizwan-ul-Haq, M.; Al-Jabr, A.M. Status of insecticide resistance in field-collected populations of Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae). Int. J. Agric. Biol. 2016, 18, 103–110. [Google Scholar] [CrossRef]
- Hussain, A.; Rizwan-ul-Haq, M.; Al-Ayedh, H.; AlJabr, A. Susceptibility and immune defence mechanisms of Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) against entomopathogenic fungal infections. Int. J. Mol. Sci. 2016, 17, 1518. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Rizwan-ul-Haq, M.; Al-Ayedh, H.; Ahmed, S.; Al-Jabr, A.M. Effect of Beauveria bassiana infection on the feeding performance and antioxidant defence of red palm weevil, Rhynchophorus ferrugineus. BioControl 2015, 60, 849–859. [Google Scholar] [CrossRef]
- Dembilio, Ó.; Quesada-Moraga, E.; Santiago-Álvarez, C.; Jacas, J.A. Potential of an indigenous strain of the entomopathogenic fungus Beauveria bassiana as a biological control agent against the red palm weevil, Rhynchophorus ferrugineus. J. Invertebr. Pathol. 2010, 104, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Dembilio, Ó.; Llácer, E.; de Martínez Altube, M.; Jacas, J.A. Field efficacy of Imidacloprid and Steinernema carpocapsae in a chitosan formulation against the red palm weevil Rhynchophorus ferrugineus (Coleoptera: Curculionidae) in Phoenix canariensis. Pest Manag. Sci. 2010, 66, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Hou, Y. Isolation and identification of bacterial strains with insecticidal activities from Rhynchophorus ferrugineus Oliver (Coleoptera: Curculionidae). J. Appl. Entomol. 2016, 140, 617–626. [Google Scholar] [CrossRef]
- Francesca, N.; Alfonzo, A.; Verde, G.L.; Settanni, L.; Lucido, P. Biological activity of Bacillus spp. evaluated on eggs and larvae of red palm weevil Rhynchophorus ferrugineus. Ann. Microbiol. 2015, 65, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Guarino, S.; Colazza, S.; Peri, E.; Bue, P.Lo.; Germanà, M.P.; Kuznetsova, T.; Gindin, G.; Soroker, V. Behaviour-modifying compounds for management of the red palm weevil (Rhynchophorus ferrugineus Oliver). Pest Manag. Sci. 2015, 71, 1605–1610. [Google Scholar] [CrossRef] [PubMed]
- Guarino, S.; Peri, E.; Lo Bue, P.; Germanà, M.P.; Colazza, S.; Anshelevich, L.; Ravid, U.; Soroker, V. Assessment of synthetic chemicals for disruption of Rhynchophorus ferrugineus response to attractant-baited traps in an urban environment. Phytoparasitica 2013, 41, 79–88. [Google Scholar] [CrossRef]
- Hoddle, M.; Al-Abbad, A.H.; El-Shafie, H.; Faleiro, J.; Sallam, A.; Hoddle, C. Assessing the impact of areawide pheromone trapping, pesticide applications, and eradication of infested date palms for Rhynchophorus ferrugineus (Coleoptera: Curculionidae) management in Al Ghowaybah, Saudi Arabia. Crop Prot. 2013, 53, 152–160. [Google Scholar] [CrossRef]
- Abbas, M.S.T.; Hanounik, S.B.; Shahdad, A.S.; AI-Bagham, S.A. Aggregation pheromone traps, a major component of IPM strategy for the red palm weevil, Rhynchophorus ferrugineus in date palms (Coleoptera: Curculionidae). J. Pest Sci. 2006, 79, 69–73. [Google Scholar] [CrossRef]
- Al-Ayedh, H.Y.; Rasool, K.G. Determination of the optimum sterilizing radiation dose for control of the red date palm weevil Rhynchophorus ferrugineus Oliv. (Coleoptera: Curculionidae). Crop Prot. 2010, 29, 1377–1380. [Google Scholar] [CrossRef]
- Paoli, F.; Dallai, R.; Cristofaro, M.; Arnone, S.; Francardi, V.; Roversi, P.F. Morphology of the male reproductive system, sperm ultrastructure and γ-irradiation of the red palm weevil Rhynchophorus ferrugineus Oliv. (Coleoptera: Dryophthoridae). Tissue Cell 2014, 46, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Mazza, G.; Inghilesi, A.F.; Stasolla, G.; Cini, A.; Cervo, R.; Benvenuti, C.; Francardi, V.; Cristofaro, M.; Arnone, S.; Roversi, P.F. Sterile Rhynchophorus ferrugineus males efficiently impair reproduction while maintaining their sexual competitiveness in a social context. J. Pest Sci. 2015, 89, 459–468. [Google Scholar] [CrossRef]
- El Naggar, S.E.M.; Mohamed, H.F.; Mahmoud, E.A. Studies on the morphology and histology of the ovary of red palm weevil female irradiated with gamma rays. J. Asia. Pac. Entomol. 2010, 13, 9–16. [Google Scholar] [CrossRef]
- Shukla, P.; Vidyasagar, P.S.P.V.; Aldosari, S.A.; Abdel-Azim, M. Antifeedant activity of three essential oils against the red palm weevil, Rhynchophorus ferrugineus. Bull. Insectol. 2012, 65, 71–76. [Google Scholar]
- Cangelosi, B.; Clematis, F.; Monroy, F.; Roversi, P.F.; Troiano, R.; Curir, P.; Lanzotti, V. Filiferol, a chalconoid analogue from Washingtonia filifera possibly involved in the defence against the red palm weevil Rhynchophorus ferrugineus Olivier. Phytochemistry 2015, 115, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Salama, H.; Ismail, I. Potential of certain natural extracts for the control of the red palm weevil, Rhynchophorus ferrugineus (Oliver). Arch. Phytopathol. Plant Prot. 2007, 40, 233–236. [Google Scholar] [CrossRef]
- Abdullah, M.A. Toxicological and histopathological studies of Boxus chinensis oil and precocene II on larvae of the red palm weevil Rynchophorus ferrugineus (Oliver) (Coleoptera: Curculionidae). Egypt. Acad. J. Biol. Sci. 2009, 2, 45–54. [Google Scholar]
- Nascimento, N.C.; Fett-Neto, A.G. Plant secondary metabolism and challenges in modifying its operation: An overview. In Plant Secondary Metabolism Engineering Methods and Applications; Fett-Neto, A.G., Ed.; Humana Press: New York, NY, USA, 2010; pp. 1–13. [Google Scholar]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.H.; Nishida, R. Methyl eugenol: Its occurrence, distribution, and role in nature, especially in relation to insect behavior and pollination. J. Insect Sci. 2012, 12. [Google Scholar] [CrossRef] [PubMed]
- Scott, I.; Jensen, H.; Scott, J.; Isman, M.; Arnason, J.; Philogène, B. Botanical insecticides for controlling agricultural pests: Piperamides and the Colorado Potato Beetle Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae). Arch. Insect Biochem. Physiol. 2003, 54, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Grant, G.; Langevin, D.; MacDonald, L. Deterring and inhibiting effects of quinolizidine alkaloids on spruce budworm (Lepidoptera: Tortricidae) oviposition. Environ. Entomol. 1998, 27, 984–992. [Google Scholar] [CrossRef]
- Koul, O. Phytochemicals and insect control: An antifeedant approach. CRC Crit. Rev. Plant Sci. 2008, 27, 1–24. [Google Scholar] [CrossRef]
- Nassar, M.; Abdulah, A. Assessment of Boxus chinensis oil and precocene II for the control of the red palm weevil, Rhychophorus ferrugineus (Oliv.) (Coleoptera-Curculionidae) and the palm beetle, Pseudophilus testaceous. J. Entomol. 2005, 2, 1–8. [Google Scholar]
- El-Bokl, M.M.; Baker, R.F.; El-Gammal, H.L.; Mahmoud, M.Z. Biological and histopathological effects of some insecticidal agents against red palm weevil Rhynchophorus ferrugineus. Egypt. Acad. J. Biol. Sci. Histol. Histochem. 2010, 1, 7–22. [Google Scholar]
- Nassar, M.; Abdullah, M. Evaluation of azadirachtin for control of the red palm weevil Rhynchophorus ferrugineus (Olivier) (Coleoptera, Curculionidae). J. Egypt. Ger. Soc. Zool. 2001, 36, 163–173. [Google Scholar]
- Abdullah, M.A.R. Identification of the biological active compounds of two natural extracts for the control of the red palm weevil, Rhynchophorus ferrugineus (Oliver) (Coleoptera: Curculionidae). Egypt Acad. J. Biol. Sci. 2009, 2, 35–44. [Google Scholar]
- Ahn, Y.J.; Kim, N.J.; Byun, S.G.; Cho, J.E.; Chung, K. Larvicidal activity of Kaempferia galanga rhizome phenylpropanoids towards three mosquito species. Pest Manag. Sci. 2008, 64, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, L.; Liu, Z.; Du, S. Identification of insecticidal constituents of the essential oil of Acorus calamus rhizomes against Liposcelis bostrychophila Badonnel. Molecules 2013, 18, 5684–5696. [Google Scholar] [CrossRef] [PubMed]
- Moreira, M.D.; Picanço, M.C.; de Barbosa, L.C.A.; Guedes, R.N.C.; de Campos, M.R.; Silva, G.A.; Martins, J.C. Plant compounds insecticide activity against coleoptera pests of stored products. Pesqui. Agropecuária Bras. 2007, 42, 909–915. [Google Scholar] [CrossRef]
- Sharma, R.; Negi, D.S.; Shiu, W.K.P.; Gibbons, S. Characterization of an insecticidal coumarin from Boenninghausenia albiflora. Phyther. Res. 2006, 20, 607–609. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Tian, M.Y.; He, Y.R.; Ahmed, S. Entomopathogenic fungi disturbed the larval growth and feeding performance of Ocinara varians (Lepidoptera: Bombycidae) larvae. Insect Sci. 2009, 16, 511–517. [Google Scholar] [CrossRef]
- Rashid, M.; Khan, R.A.; Zhang, Y.L. Over-expression of cytochrome P450s in Helicoverpa armigera in response to bioinsecticide, cantharidin. Int. J. Agric. Biol. 2013, 15, 993–997. [Google Scholar]
- Halon, E.; Eakteiman, G.; Moshitzky, P.; Elbaz, M.; Alon, M.; Pavlidi, N.; Vontas, J.; Morin, S. Only a minority of broad-range detoxification genes respond to a variety of phytotoxins in generalist Bemisia tabaci species. Sci. Rep. 2015, 5, 17975. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.-G.; Wang, S.; Dou, T.-T.; Liu, S.; Li, M.-Y.; Hua, R.-M.; Li, S.-G.; Lin, H.-F. Aphicidal activity of Illicium verum fruit extracts and their effects on the Acetylcholinesterase and Glutathione S-transferases activities in Myzus persicae (Hemiptera: Aphididae). J. Insect Sci. 2016, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Francis, F.; Vanhaelen, N.; Haubruge, E. Glutathione S-transferases in the adaptation to plant secondary metabolites in the Myzus persicae aphid. Arch. Insect Biochem. Physiol. 2005, 58, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Russell, R.M.; Robertson, J.L.; Savin, N.E. POLO: A new computer program for Probit analysis. Bull. Entomol. Soc. Am. 1977, 23, 209–213. [Google Scholar] [CrossRef]
- Statistix Statistix 8.1: Analytical Software 2003; Analytical Software: Tallahassee, FL, USA; Available online: http://statistix.com (accessed on 19 January 2017).
- SAS Institute. SAS User’s Guide: Statistics; SAS Institute, Cary 2000; SAS: Cary, NC, USA; Available online: www.sas.com (accessed on 19 January 2017).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.




{kind=link}
{kind=link}
{kind=link}
| Treatments | LD50 (g/L) | 95% CL | χ2 | Slope ± SE |
|---|---|---|---|---|
| Coumarin 1 | 0.672 | 0.580–0.778 | 4.51 | 1.91 ± 0.25 |
| Ethyl cinnamate 2 | 6.974 | 6.113–7.955 | 5.60 | 2.07 ± 0.25 |
| Diniconazole 2 | 9.756 | 8.531–11.158 | 4.42 | 1.99 ± 0.25 |
| Isoeugenol 2 | 11.560 | 10.080–13.270 | 4.50 | 1.98 ± 0.25 |
| Coumarin 6 2 | 13.330 | 11.580–15.350 | 1.57 | 1.87 ± 0.25 |
| α-Asarone 2 | 17.670 | 15.040–20.750 | 5.94 | 1.80 ± 0.25 |
| Eugenol 2 | 23.260 | 18.020–30.030 | 1.81 | 1.43 ± 0.25 |
| Coniferyl aldehyde 2 | 27.010 | 19.740–36.970 | 1.58 | 1.34 ± 0.25 |
| Rosmarinic acid 3 | n/a | n/a | n/a | n/a |
| Methyl isoeugenol 3 | n/a | n/a | n/a | n/a |
| Methyl eugenol 3 | n/a | n/a | n/a | n/a |
| Treatments | AD | ECI | ECD |
|---|---|---|---|
| Coumarin | 74.01 ± 0.35 a | 05.75 ± 0.53 f | 07.76 ± 0.71 h |
| Ethyl cinnamate | 60.02 ± 0.15 b | 15.01 ± 0.17 e | 25.01 ± 0.30 g |
| Diniconazole | 59.02 ± 0.44 bc | 16.01 ± 0.11 d | 27.13 ± 0.20 f |
| Isoeugenol | 58.06 ± 0.50 cd | 16.88 ± 0.24 c | 29.09 ± 0.62 e |
| Coumarin 6 | 57.71 ± 0.32 de | 17.34 ± 0.21 bc | 30.05 ± 0.53 de |
| α-Asarone | 57.17 ± 0.49 de | 17.52 ± 0.19 bc | 30.66 ± 0.60 cde |
| Eugenol | 56.72 ± 0.40 ef | 17.69 ± 0.30 b | 31.20 ± 0.73 cd |
| Coniferyl aldehyde | 55.92 ± 0.25 fg | 17.71 ± 0.27 b | 31.68 ± 0.62 cd |
| Rosmarinic acid | 55.66 ± 0.33 g | 17.91 ± 0.10 b | 32.19 ± 0.31 c |
| Methyl isoeugenol | 55.34 ± 0.45 gh | 18.81 ± 0.40 a | 34.02 ± 0.99 b |
| Methyl eugenol | 54.33 ± 0.36 hi | 19.03 ± 0.30 a | 35.04 ± 0.74 ab |
| Control | 53.45 ± 0.28 i | 19.19 ± 0.21 a | 35.91 ± 0.58 a |
| Target Gene | Accession No | Amplicon Size | Primer 5′–3′ (Forward and Reverse) |
|---|---|---|---|
| Detoxification | |||
| Cytochrome P450 | KT748789 | 118 bp | TGGAGAAACACCCGCAAGAA CGGCGATTTTGCCTACCAAG |
| Glutathione S-transferase | KR902496 | 92 bp | ATAGCCAACCACCACTGTCG CGTTCCTCTTGCCGCTAGTT |
| Esterase | KT748822 | 70 bp | ACCTACAAGAATCCGACGCC ACTCCGAAACTTTGGGCCAT |
| Housekeeping | |||
| beta-Actin | KM438516 | 129 bp | AAAGGTTCCGTTGCCCTGAA TGGCGTACAAGTCCTTCCTG |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
AlJabr, A.M.; Hussain, A.; Rizwan-ul-Haq, M.; Al-Ayedh, H. Toxicity of Plant Secondary Metabolites Modulating Detoxification Genes Expression for Natural Red Palm Weevil Pesticide Development. Molecules 2017, 22, 169. https://doi.org/10.3390/molecules22010169
AlJabr AM, Hussain A, Rizwan-ul-Haq M, Al-Ayedh H. Toxicity of Plant Secondary Metabolites Modulating Detoxification Genes Expression for Natural Red Palm Weevil Pesticide Development. Molecules. 2017; 22(1):169. https://doi.org/10.3390/molecules22010169
Chicago/Turabian StyleAlJabr, Ahmed Mohammed, Abid Hussain, Muhammad Rizwan-ul-Haq, and Hassan Al-Ayedh. 2017. "Toxicity of Plant Secondary Metabolites Modulating Detoxification Genes Expression for Natural Red Palm Weevil Pesticide Development" Molecules 22, no. 1: 169. https://doi.org/10.3390/molecules22010169