Selenium Cycling Across Soil-Plant-Atmosphere Interfaces: A Critical Review

Abstract
:1. Introduction
2. Selenium in Crops
2.1. Biofortification
2.1.1. Speciation of Se in Crops
2.1.2. Application of Se to Soils
2.2. Genetic Engineering
Using Naturally-Enriched Se Materials
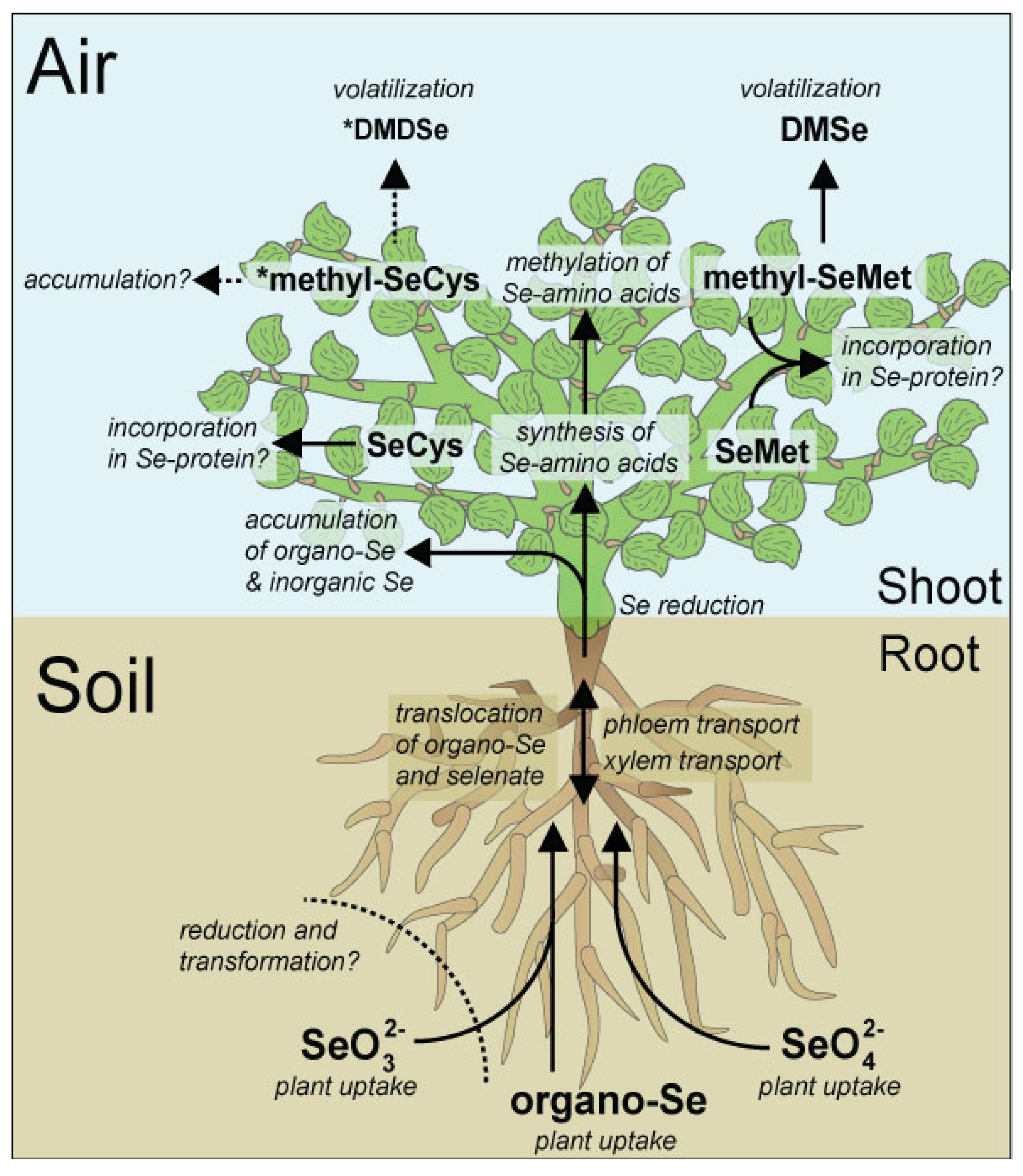
3. Sources and Sinks of Soil Selenium

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject/Scope | Year | Reference |
|---|---|---|
| Overview of analytical methods, bioaccumulation, and effects of Se in agricultural environments. | 1984 | [43] |
| Overview of the state of knowledge of Se occurrence, effects, uptake, etc. in agricultural environments. | 1989 | [44] |
| Comparison between Se and S biogeochemical atmospheric cycling and discussion of atmospheric Se fluxes. | 1990 | [45] |
| Selenium speciation, partitioning, and volatilization in wetlands. | 1993 | [46] |
| Occurrence of Se in natural and environmental waters, incl. a review of analytical methods and speciation. | 1997 | [47] |
| Physiology and biochemistry of Se accumulator and non-Se accumulator plants, incl. Se uptake, assimilation, incorporation into proteins and volatilization. | 2000 | [34] |
| Review of solubility and acid/base equilibration constants for various Se species. | 2001 | [48] |
| Gives an overview of biomethylation of Se and tellurium (Te) by Microorganisms and plants, incl. a review of analytical techniques and proposed biomethylation mechanisms. | 2003 | [49] |
| A review of measurement techniques to investigate Se isotope fractionation and current understanding of Se isotope dynamics. | 2004 | [50] |
| Reviews Se impacts on aquatic ecosystems and food chains. The interaction of Se with other elements and effects of Se on survival and growth of fish are also discussed. | 2004 | [12] |
| Reviews studies on Se accumulation in sediments and waters due to agricultural drainage as well as effects of Se toxicity on wildlife. | 2004 | [51] |
| Review of Se concentrations and speciation in coal and of genetic types of Se-accumulations in coal. | 2006 | [52] |
| Major aspects of atmospheric Se and its natural and anthropogenic sources. | 2007 | [40] |
| Mechanistic evaluation of processes governing Se cycling and bioavailability. | 2009 | [13] |
| Covers many facets of biofortification. | 2009 | [17] |
| Covers biofortification of agricultural crops with different macro- and micronutrients. Reviews aspects of soil, physiology, agronomic, and genetic approaches to biofortify food crops with different mineral elements. | 2009 | [53] |
| Reviews key developments in the understanding of Se in higher plants and advances in genetic engineering of Se metabolism. | 2009 | [4] |
| Reviews transgenic approaches for enhancement of plant Se accumulation, tolerance and volatilization. | 2009 | [35] |
| Review of knowledge of the role of Se in environmental sciences and discusses potential treatment options of Se contaminated waste streams. | 2009 | [54] |
| Review of plant Se metabolism and discussion of new insights into plant Se tolerance and hyperaccumulation mechanisms. | 2010 | [55] |
| Summarizes the knowledge on important organo-Se and Te species in environmental compartments, and identifies gaps and uncertainties in past and current analytical methodology. | 2010 | [14] |
| Discusses how Se phytoremediation may have ecological implications, particularly how high Se levels in plants affect herbivory. | 2011 | [56] |
| Review of how Se hyperaccumulators may profoundly affect their local ecosystem, by altering soil Se speciation and distribution and by selecting against Se-sensitive and for Se-resistant ecological partners. | 2012 | [57] |
| Overview of major aspects of volcanic–derived Se, with focus on processes in soils and aquifers. | 2012 | [41] |
| Discusses the role and importance of Se as selenoproteins in supplementing human diet. | 2013 | [24] |
| Se availability in semiarid soils and discusses how micronutrients can be effectively managed through the recycling of organic matter. | 2013 | [58] |
| Reviews studies on sorption of sorption, speciation and bioavailability of Se in non-flooded and wetland soils, incl. impact of redox/pH conditions, metal oxides, and organic matter. | 2014 | [59] |
| Summarizes what is known about plant metabolism of Se, and how plants may be used to clean up Se from agricultural drainage water. | 2014 | [60] |
| Summarizes the abundance and forms of Se in rocks, soils, sediments and natural waters and pathways and of Se in the natural environment | 2014 | [42] |
| Reviews the biogeochemistry of Se in the natural environment, in terms of variation of speciation with pH, redox conditions, sorption and interactions with natural organic matter. | 2015 | [61] |
| Overview of the role of Se-respiring bacteria (SeRB) in the biological Se cycle, their ecological role as well as Se biomineralization mechanisms and environmental biotechnological applications. | 2015 | [62] |
| Reviews what is known about plant metabolism of Se, molecular mechanisms of Se tolerance and (hyper) accumulation, achievements in plant genetic engineering of Se metabolism, and briefly touches on evolutionary and ecological aspects of plant Se accumulation. | 2015 | [63] |
| Discusses the function of Se in plant and human nutrition and the progress in the genetic engineering of Se metabolism to increase the levels and bioavailability of Se in food crops. | 2015 | [64] |

3.1. Geogenic Sources of Soil Selenium
3.2. Atmospheric Sources of Soil Selenium
Marine-Derived Atmospheric Selenium
3.3. Atmospheric Selenium Deposition
3.4. Anthropogenic versus Natural Sources of Atmospheric Selenium
Quantifying Se Sources and Sinks in Soils
4. The Mobility and Bioavailability of Soil Se
4.1. Factors Affecting Se Mobility in Soils
The Role of Organic Matter in Se Retention by Soils
4.2. Soil Factors Affecting Uptake of Se by Plants
4.2.1. The Role of Soil OM in Plant Se Uptake

4.2.2. The Role of Combined Soil Properties in Plant Se Uptake

5. Mechanisms of Se Uptake and Metabolism
5.1. Selenium Uptake by Plants
5.1.1. Uptake of inorganic Se
5.1.2. Uptake of Organic Se
5.2. Uptake of Inorganic and Organic Se by Other Organisms

Selenium Metabolism
6. Selenium Hyperaccumulation by Plants
6.1. Physological and Ecological Benefits
6.2. Mechanisms of Plant Se Hyperaccumulation
6.3. Ecological Impacts of Hyperaccumulators on Seleniferous Areas
7. Biomethylation of Se
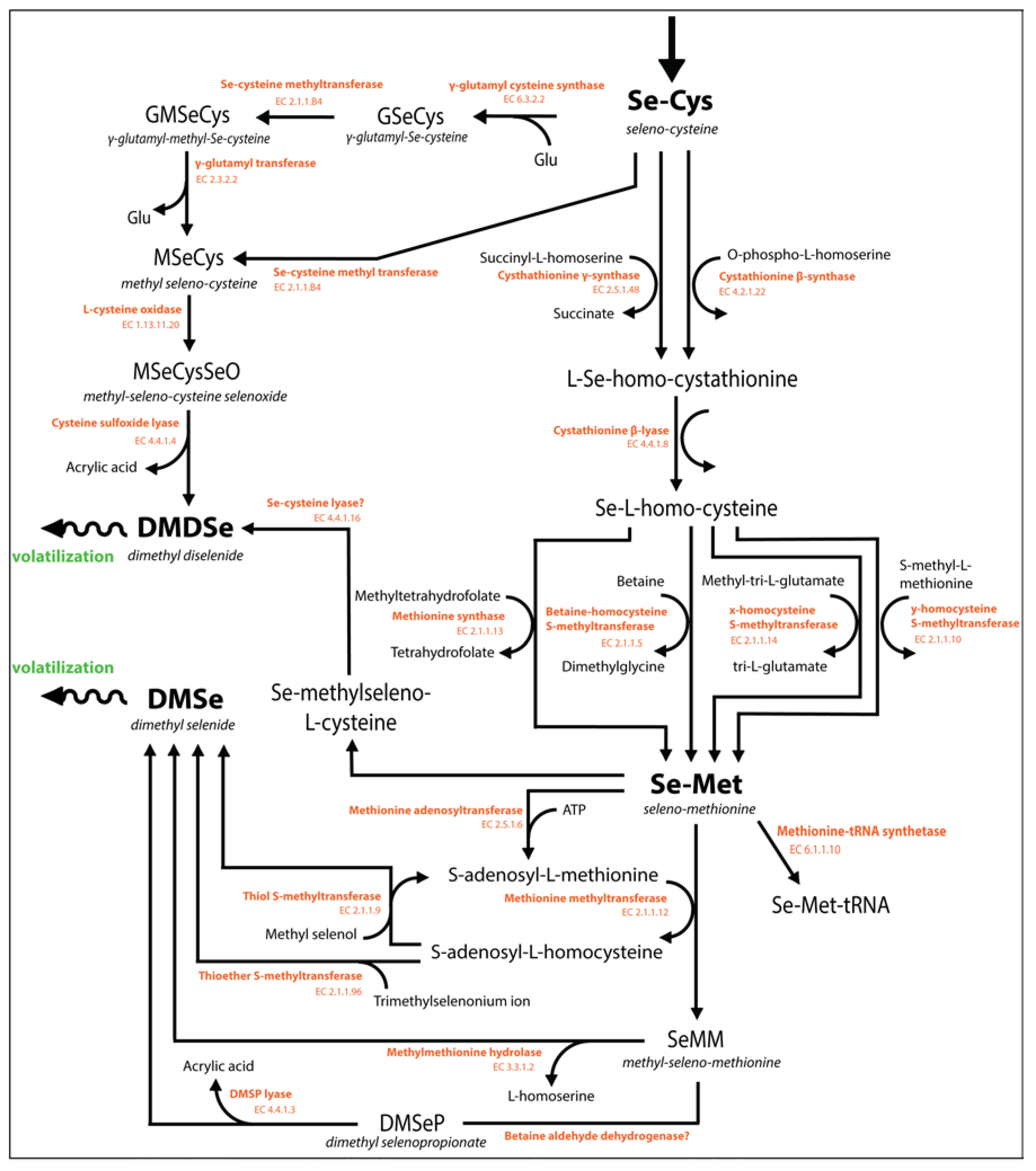
7.1. The Mechanism of Se Biomethylation

7.1.1. Selenium Methylation by Plants
7.1.2. Formation of DMSe versus DMDSe
7.2. The Occurrence of Methylated Se in Soils, Water and Air
7.2.1. Measurements of Biogenic Se Emissions
| Reference | Production of Volatile Se | Experiment | Amendment | Environment | Se Content |
|---|---|---|---|---|---|
| (Lin, Cervinka et al. 2002) [266] | 11–155 μg Se·m−2·d−1 | Field | No | Soil | 4.0 mg·kg−1 |
| (Bañuelos and Lin 2007) [37] | 25 μg Se·m−2·d−1 | Field | No | Soil | 4.78 mg·kg−1 |
| (Bañuelos, Lin et al. 2005) [267] | 3–56 μg Se·m−2·d−1 | Field | No | Soil | 3–8.2 mg·kg−1 |
| (Wu and Huang 1991) [270] | 180 μg Se·m−2·d−1 | Field | No | Soil | 1 mg·L−1 |
| (Dungan, Stork et al. 2000) [271] | 17–72 mg Se·m−2·h−1 | Field | No | Soil | 0.14 mg·kg−1 |
| (Zawislanski 1996) [272] | 20–200 μg Se·m−2·d−1 | Field | No | Soil | 50–180 mg·kg−1 |
| (Lin, Hansen et al. 1999) [273] | 117–125 μg Se·m−2·d−1 | Field | No | Soil | 4–8 mg·kg−1 |
| (Bañuelos and Lin 2007) [37] | 114–434 μg Se·m−2·d−1 | Field | Yes | Soil | 4.78 mg·kg−1 |
| (Frankenberger W.T. Jr. 1995) [265] | 72–1300 μg Se·m−2·d−1 | Field | Yes | Soil | 11.4 mg·kg−1 |
| (Bañuelos, Terry et al. 2005) [36] | 4–13.4 μg Se·m−2·d−1 | Laboratory | No | Soil | 0.15–13 mg·kg−1 |
| (Wu and Huang 1991) [270] | 65 μg Se·gsoil−1·d−1 * | Laboratory | No | Soil | 1 mg·L−1 |
| (Martens and Suarez 2003) [274] | <1 μg Se·kgsoil−1·d−1 | Laboratory | No | Soil | 10 mg·kg−1 |
| (Moreno-Jiménez, Clemente et al. 2013) [275] | 4–214 μg Se·kgsoil−1·d−1 | Laboratory | Yes | Soil | 68 mg·kg−1 |
| (Zieve and Peterson 1981) [276] | 2 μg Se·kgsoil−1·d−1 | Laboratory | Yes | Soil | 5 mg·kg−1 |
| (Stork, Jury et al. 1999) [262] | <700 μg Se·kgsoil−1·d−1 | Laboratory | Yes | Soil | 18.3–0.14 mg·kg−1 |
| (Frankenberger and Karlson 1989) [277] | <1.1 mg Se·gsoil−1·d−1 | Laboratory | Yes | Soil | 60 g·kg−1 |
| (Dhillon, Dhillon et al. 2010) [278] | <32 ng Se·kgsoil−1·d−1 | Laboratory | Yes | Soil | 1.7–31 mg·kg−1 |
| (Karlson and Frankenberger 1988) [263] | <677 μg Se·kgsoil−1·d−1 | Laboratory | Yes | Soil | 100 mg·kg−1 |
| (Vriens, Lenz et al. 2014) [94] | 0.11–0.12 μg Se·m−2·d−1 | Field | No | Wetland | 2.0 ± 0.4 mg·kg−1 |
| (Vriens, Ammann et al. 2014) [261] | 190–210 ng Se·m−2·d−1 | Field | No | Wetland | 2.0 ± 0.4 mg·kg−1 |
| (Hansen, Duda et al. 1998) [279] | 25–190 μg Se·m−2·d−1 | Field | No | Wetland | 4–31 μg·L−1 |
| (Gao, Tanji et al. 2003)[280] | 44–285 μg Se·m−2·d−1 | Field | No | Wetland | 10 μg·L−1 |
| (Zhang and Moore 1997) [281] | 0.1–60 μg Se·m−2·d−1 | Laboratory | Yes | Wetland | 2–11 μg·L−1 |
| (Thompson-Eagle and Frankenberger 1991) [282,283] | 0.1–10 μg Se·Lwater−1·d−1 | Laboratory | Yes | Wetland | 0.02–102 mg·L−1 |
| (Thompson-Eagle and Frankenberger 1990) [282,283] | 0.55–6.9 μg Se·Lwater−1·d−1 | Laboratory | Yes | Wetland | 14–2000 μg·L−1 |
| (Thompson-Eagle and Frankenberger 1990) [282,283] | <18 μg Se·Lwater−1·d−1 | Laboratory | Yes | Wetland | 0.7–2.9 mg·L−1 |
| (Calderone, Frankenberger et al. 1990) [284] | <25 μg Se·kgsediment−1·d−1 | Laboratory | Yes | Sediment | <40.7 mg·kg−1 |
| (Diaz, Johnson et al. 2008) [269] | <1.8 μg Se·m−2·d−1 | Field | No | Lake | 6.3 nM |
| (Amouroux and Donard 1997) [258] | 0.002–0.5 μg Se·m−2·d−1 | Field | No | Estuary | 0.06–1.12 nM |
| (Terry, Carlson et al. 1992) [268] | 15–350 μg Se·m−2·d−1 (m2 leaf area) * 0.2–2.5 mg Se·kgplant−1·d−1 | Laboratory | Yes | Plant leaf | < 20 μM |
| (Amouroux and Donard 1996) [96] | 0.002–0.23 μg Se·m−2·d−1 | Field | No | Marine | ~1 nM |
| (Fan, Higashi et al. 1998) [285] | <0.75 μg Se·Lwater−1·d−1 | Laboratory | Yes | Marine | Up to 1 mg·L−1 |
7.2.2. Limitations of Emission Data
8. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Combs, G.F. Selenium in global food systems. Brit. J. Nutr. 2001, 85, 517–547. [Google Scholar] [CrossRef] [PubMed]
- Finley, J.W. Selenium accumulation in plant foods. Nutr. Rev. 2005, 63, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Pilon-Smits, E.A.; Zhao, F.J.; Williams, P.N.; Meharg, A.A. Selenium in higher plants: Understanding mechanisms for biofortification and phytoremediation. Trends Plant Sci. 2009, 14, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Cone, J.E.; Del Río, R.M.; Davis, J.N.; Stadtman, T.C. Chemical characterization of the selenoprotein component of clostridial glycine reductase: Identification of selenocysteine as the organoselenium moiety. Proc. Natl. Acad. Sci. USA 1976, 73, 2659–2663. [Google Scholar] [CrossRef] [PubMed]
- Fairweather-Tait, S.J.; Collings, R.; Hurst, R. Selenium bioavailability: Current knowledge and future research requirements. Am. J. Clin. Nutr. 2010, 91, 1484S–1491S. [Google Scholar] [CrossRef] [PubMed]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigó, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef] [PubMed]
- Castellano, S.; Gladyshev, V.N.; Guigo, R.; Berry, M.J. SelenoDB 1.0: A database of selenoprotein genes, proteins and SECIS elements. Nucleic Acids Res. 2008, 36, (Suppl. 1). D332–D338. [Google Scholar] [CrossRef] [PubMed]
- Fairweather-Tait, S.J.; Bao, Y.; Broadley, M.R.; Collings, R.; Ford, D.; Hesketh, J.R.; Hurst, R. Selenium in human health and disease. Antioxid. Redox Signal. 2011, 14, 1337–1383. [Google Scholar] [CrossRef] [PubMed]
- Steinnes, E. Soils and geomedicine. Environ. Geochem. Hlth. 2009, 31, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P.; Infante, H.G.; Sargent, M. Food-chain selenium and human health: Spotlight on speciation. Br. J. Nutr. 2008, 100, 238–253. [Google Scholar] [PubMed]
- Hamilton, S.J. Review of selenium toxicity in the aquatic food chain. Sci. Total Environ. 2004, 326, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Martinez, A.; Charlet, L. Selenium environmental cycling and bioavailability: A structural chemist point of view. Rev. Environ. Sci. Biotechnol. 2009, 8, 81–110. [Google Scholar] [CrossRef]
- Wallschlaeger, D.; Feldmann, J. Formation, occurrence, significance, and analysis of organoselenium and organotellurium compounds in the environment. In Organometallics in Environment and Toxicology; Sigel, A., Ed.; Royal Society of Chemistry: Cambridge, UK, 2010; Volume 7, pp. 319–364. [Google Scholar]
- Zwolak, I.; Zaporowska, H. Selenium interactions and toxicity: A review. Cell Biol. Toxicol. 2012, 28, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, J.E. Selenium World Atlas: Updated Edition; Selenium-Tellurium Dev. Assoc.: Grimbergen, Belgium, 2002. [Google Scholar]
- Bañuelos, G.S.; Lin, Z.-Q. Development and Use of Biofortified Agricultural Products; CRC Press: Boca Raton, FL, USA, 2009; p. 297. [Google Scholar]
- Broadley, M.R.; White, P.J.; Bryson, R.J.; Meacham, M.C.; Bowen, H.C.; Johnson, S.E.; Hawkesford, M.J.; McGrath, S.P.; Zhao, F.J.; Breward, N.; et al. Biofortification of UK food crops with selenium. Proc. Nutr. Soc. 2006, 65, 169–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiry, C.; Schneider, Y-J.; Pussemier, L.; de Temmerman, L.; Ruttens, A. Selenium bioaccessibility and bioavailability in Se-enriched food supplements. Biol. Trace Elem. Res. 2013, 152, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Poblaciones, M.J.; Rodrigo, S.; Santamaría, O.; Chen, Y.; McGrath, S.P. Agronomic selenium biofortification in triticum durum under mediterranean conditions: From grain to cooked pasta. Food Chem. 2014, 146, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Kápolna, E.; Hillestrøm, P.R.; Laursen, K.H.; Husted, S.; Larsen, E.H. Effect of foliar application of selenium on its uptake and speciation in carrot. Food Chem. 2009, 115, 1357–1363. [Google Scholar] [CrossRef]
- Kápolna, E.; Laursen, K.H.; Husted, S.; Larsen, E.H. Bio-fortification and isotopic labelling of Se metabolites in onions and carrots following foliar application of Se and 77Se. Food Chem. 2012, 133, 650–657. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Arroyo, I.; Pickering, I.J.; Yang, S.I.; Freeman, J.L. Selenium biofortification of broccoli and carrots grown in soil amended with Se-enriched hyperaccumulator Stanleya pinnata. Food Chem. 2014. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Blazejak, S. Selenium: Significance, and outlook for supplementation. Nutrition 2013, 29, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.; Ou, B.; Liang, Y.; Yeh, F. Growth inhibition and antioxidative status induced by selenium enriched broccoli extract and selenocompounds in DNA mismach repair-deficient human colon cancer cells. Food Chem. 2013, 139, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Alfthan, G.P.; Aspila, P.; Ekhyolm, M.; Hartikainen., H.; Hero, H.; Hietaniemi, V.; Root, T.; Salminen, P.; Venäläinen, E.-R.; Aro, A. Nationwide supplementation of sodium selenate to commercial fertilizers: History and 25 year results from the Finnish selenium monitoring program. In Combating Micronutrient Deficiencies: Food-Based Approaches; Thompson, B., Amoroso, L., Eds.; FAO/CAB International: Rome, Italy, 2011; pp. 312–337. [Google Scholar]
- Hartikainen, H. Biogeochemistry of selenium and its impact on food chain quality and human health. J. Trace Elem. Med. Biol. 2005, 18, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Lyons, G. Selenium in cereals: Improving the efficiency of agronomic biofortification in the UK. Plant Soil 2010, 332, 1–4. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Curtin, D.; Hanson, R.; Lindley, T.N.; Butler, R.C. Selenium concentration in wheat grain as influenced by method, rate, and timing of sodium selenate application. N. Z. J. Crop Hort. 2006, 34, 324–339. [Google Scholar] [CrossRef]
- Zhao, C.Y.; Ren, J.H.; Xue, C.Z.; Lin, E. Study on the relationship between soil Se and plant Se uptake. Plant Soil 2005, 277, 197–206. [Google Scholar] [CrossRef]
- Ohlendorf, H.M.; Hoffman, D.J.; Saiki, M.K.; Aldrich, T.W. Embryonic mortality and abnormalities of aquatic birds: Apparent impacts of selenium from irrigation drain-water. Sci. Total Environ. 1986, 52, 49–63. [Google Scholar] [CrossRef]
- Freeman, J.L.; Tamaoki, M.; Stushnoff, C.; Quinn, C.F.; Cappa, J.J.; Devonshire, J.; Fakra, S.; Marcus, M.A.; McGrath, S.; van Hoewyk, D.; et al. Molecular mechanisms of selenium tolerance and hyperaccumulation in Stanleya pinnata. Plant Physiol. 2010, 153, 1630–1652. [Google Scholar] [CrossRef] [PubMed]
- Terry, N.; Zayed, A.M.; de Souza, M.P.; Tarun, A.S. Selenium in higher plants. Ann. Rev. Plant Physiol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.H.; LeDuc, D.L. Phytoremediation of selenium using transgenic plants. Curr. Opin. Biotechnol. 2009, 20, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos, G.S.; Terry, N.; Leduc, D.L.; Pilon-Smits, E.A.H.; Mackey, B. Field trial of transgenic Indian mustard plants shows enhanced phytoremediation of selenium-contaminated sediment. Environ. Sci. Technol. 2005, 39, 1771–1777. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Lin, Z.-Q. Acceleration of selenium volatilization in seleniferous agricultural drainage sediments amended with methionine and casein. Environ. Pollut. 2007, 150, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos, G.S. Irrigation of broccoli and canola with boron and selenium laden effluent. J. Environ. Qual. 2002, 31, 1802–1808. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos, G.S.; Freeman, J.L. Natural biofortification of vegatables. Food Chem. 2015. in preparation. [Google Scholar]
- Wen, H.; Carignan, J. Reviews on atmospheric selenium: Emissions, speciation and fate. Atmos. Environ. 2007, 41, 7151–7165. [Google Scholar] [CrossRef]
- Floor, G.H.; Román-Ross, G. Selenium in volcanic environments: A review. Appl. Geochem. 2012, 27, 517–531. [Google Scholar] [CrossRef]
- Plant, J.A.; Bone, J.; Voulvoulis, N.; Kinniburgh, D.G.; Smedley, P.L.; Fordyce, F.M.; Klinck, B. Arsenic and Selenium. In Treatise on Geochemistry, 2nd ed.; Turekian, K.H., Holland, H.D., Eds.; Elsevier: Oxford, UK, 2014; pp. 13–57. [Google Scholar]
- Gissel-Nielsen, G.; Gupta, U.C.; Lamand, M.; Westermarck, T. Selenium in soils and plants and its importance in livestock and human nutrition. Adv. Agron. 1984, 37, 397–460. [Google Scholar]
- Jacobs, L.W. (Ed.) Selenium in Agriculture and the Environment; Soil Science Society of America: Madison, WI, USA, 1989; p. 233.
- Ross, H. Biogeochemical cycling of atmospheric selenium. In Metal Speciation in the Environment; Broekaert, J.A.C., Güçer, Ş., Adams, F., Eds.; Springer: Berlin/Heidelberg, Germany, 1990; Volume 23, pp. 523–543. [Google Scholar]
- Masscheleyn, P.H.; Patrick, W.H. Biogeochemical processes affecting selenium cycling in wetlands. Environ. Toxicol. Chem. 1993, 12, 2235–2243. [Google Scholar] [CrossRef]
- Conde, J.E.; Alaejos, M.S. Selenium concentrations in natural and environmental waters. Chem. Rev. 1997, 97, 1979–2003. [Google Scholar] [CrossRef] [PubMed]
- Séby, F.; Potin-Gautier, M.; Giffaut, E.; Borge, G.; Donard, O.F.X. A critical review of thermodynamic data for selenium species at 25 °C. Chem. Geol. 2001, 171, 173–194. [Google Scholar] [CrossRef]
- Chasteen, T.G.; Bentley, R. Biomethylation of selenium and tellurium: microorganisms and plants. Chem. Rev. 2003, 103, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.M. A review of mass-dependent fractionation of selenium isotopes and implications for other heavy stable isotopes. Chem. Geol. 2004, 204, 201–214. [Google Scholar] [CrossRef]
- Wu, L. Review of 15 years of research on ecotoxicology and remediation of land contaminated by agricultural drainage sediment rich in selenium. Ecotoxicol. Environ. Saf. 2004, 57, 257–269. [Google Scholar] [CrossRef]
- Yudovich, Y.E.; Ketris, M.P. Selenium in coal: A review. Int. J. Coal Geol. 2006, 67, 112–126. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Lenz, M.; Lens, P.N. The essential toxin: The changing perception of selenium in environmental sciences. Sci. Total Environ. 2009, 407, 3620–3633. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.H.; Quinn, C.F. Selenium metabolism in plants. In Cell Biology of Metals and Nutrients; Hell, R., Mendel, R.-R., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 17, pp. 225–241. [Google Scholar]
- Quinn, Q.F.; Lindblom, S.D.; Pilon-Smits, E.A.H. Ecological aspects of selenium phytoremediation. In Phytoremediation: Processes, Characteristics, and Applications; Columbus, F., Ed.; Nova Publishers: Hauppauge, USA, 2011; pp. 651–666. [Google Scholar]
- El-Mehdawi, A.F.; Pilon-Smits, E.A.H. Ecological aspects of plant selenium hyperaccumulation. Plant Biol. 2012, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Garcia Moreno, R.; Burdock, R.; Díaz Álvarez, M.C.; Crawford, J.W. Managing the selenium content in soils in semiarid environments through the recycling of organic matter. Appl. Environ. Soil Sci. 2013, 2013, 1–10. [Google Scholar] [CrossRef]
- Nakamaru, Y.M.; Altansuvd, J. Speciation and bioavailability of selenium and antimony in non-flooded and wetland soils: A review. Chemosphere 2014, 111, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.H.; Bañuelos, G.S.; Parker, D.R. Uptake, Metabolism, and Volatilization of Selenium by Terrestrial Plants. In Salinity and Drainage in San Joaquin Valley; Chang, A.C., Brawer Silva, D., Eds.; Science, Technology, and Policy, Global Issues in Water Policy; Springer Science + Business Media: Dordrecht, The Netherlands, 2014; Volume 5, pp. 147–164. [Google Scholar]
- Sharma, V.K.; McDonald, T.J.; Sohn, M.; Anquandah, G.A.K.; Pettine, M.; Zboril, R. Biogeochemistry of selenium. A review. Environ. Chem. Lett. 2015, 13, 49–58. [Google Scholar] [CrossRef]
- Nancharaiah, Y.V.; Lens, P.N.L. Ecology and biotechnology of selenium-respiring bacteria. Microbiol. Mol. Biol. Rev. 2015, 79, 61–80. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.H. Selenium in plants. In Progress in Botany; Luettge, U., Ed.; Springer: Heidelberg, Germany, 2015; pp. 93–107. [Google Scholar]
- Malagoli, M.; Schiavon, M.; Dall’Acqua, S.; Pilon-Smits, E.A.H. Implications of selenium biofortification in food nutritional quality. Front. Plant Sci. 2015, in press. [Google Scholar] [CrossRef] [PubMed]
- Vine, J.D.; Tourtelot, E.B. Geochemistry of black shale deposit—A summary report. Econ. Geol. 1970, 65, 253–272. [Google Scholar] [CrossRef]
- Vine, J.D. Element Distribution in Some Paleozoic Black Shales and Associated Rocks: USGS Bulletin 1214-G; BiblioGov: Washington, USA, 2013; p. 40. [Google Scholar]
- Kim, J.H. Geochemistry and genesis of the Guryongsan (Ogcheon) uraniferous black slates. J. Korean Inst. Min. Geol. 1989, 22, 35–63. [Google Scholar]
- Webb, J.S.; Thornton, I.; Fletcher, W.K. Seleniferous soils in parts of England and Wales. Nature 1966, 211, 327. [Google Scholar] [CrossRef]
- Rogers, P.A.M.; Arora, S.P.; Fleming, G.A.; Crinion, R.A.P.; McLaughlin, J.G. Selenium toxicity in farm animals: Treatment and prevention. Ir. Vet. J. 1990, 43, 151–153. [Google Scholar]
- Thornton, I.; Kinniburgh, D.G.; Pullen, G.; Smith, C.A. Geochemical aspects of selenium in British soils and implications to animal health. In Trace Substances in Environmental Health; Hemphill, D.D., Ed.; University of Missouri: Columbia, DC, USA, 1983; Volume XVII, pp. 391–398. [Google Scholar]
- Wen, H.J.; Qiu, Y.Z. Organic and inorganic occurrence of selenium in Laerma Se-Au deposit. Sci. China Ser. D 1999, 42, 662–669. [Google Scholar] [CrossRef]
- Zhang, G.D.; Ge, X.L.; Zhang, Y.L.; Luo, D.H.; Liu, X.P.; Fordyce, F.M.; Mao, D.; Su, H. Selenium geological and geochemical environmental background in Enshi, Hubei. Acta Geosci. Sin. 1998, 19, 59–67, (In Chinese with English Abstract). [Google Scholar]
- Shao, S.X.; Zheng, B.S. The biogeochemistry of selenium in Sunan grassland, Gansu, Northwest China, casts doubt on the belief that Marco Polo reported selenosis for the first time in history. Environ. Geochem. Hlth 2008, 30, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Presser, T.S.; Sylvester, M.A.; Low, W.H. Bioaccumulation of selenium from natural geologic sources in western states and its potential consequences. Environ. Manag. 1994, 18, 423. [Google Scholar] [CrossRef]
- Tuttle, M.L.W.; Fahy, J.W.; Elliott, J.G.; Grauch, R.I.; Stillings, L.L. Contaminants from Cretaceous black shale: I. Natural weathering processes controlling contaminant cycling in Mancos Shale, southwestern United States, with emphasis on salinity and selenium. Appl. Geochem. 2014, 46, 57–71. [Google Scholar] [CrossRef]
- Kulp, T.R.; Pratt, L.M. Speciation and weathering of selenium in upper cretaceous chalk and shale from South Dakota and Wyoming, USA. Geochim. Cosmochim. Acta 2004, 68, 3687–3701. [Google Scholar] [CrossRef]
- Presser, T.S.; Swain, W.C. Geochemical evidence for Se mobilization by the weathering of pyritic shale, San Joaquin Valley, California, USA. Appl. Geochem. 1990, 5, 703–717. [Google Scholar] [CrossRef]
- Seiler, R.L.; Skorupa, J.P.; Peltz, L.A. Areas Susceptible to Irrigation-Induced Selenium Contamination of Water and Biota in the Western United States; U.S. Geological Survey Circular 1180: Carson City, NV, USA, 1999; p. 36. [Google Scholar]
- Ohlendorf, H.M. The birds of Kesterson Reservoir: A historical perspective. Aquat. Toxicol. 2002, 57, 1–10. [Google Scholar] [CrossRef]
- Hanson, B.R. A master plan for drainage in the San Joaquin Valley. Calif. Agric. 1982, 36, 9–11. [Google Scholar]
- Luo, K.L.; Xu, L.R.; Tan, J.A.; Wang, D.H.; Xiang, L.H. Selenium source in the selenosis area of the Daba region, South Qinling Mountain, China. Environ. Geol. 2004, 45, 426–432. [Google Scholar]
- Li, Y.H.; Wang, W.; Luo, K.L.; Li, H.R. Environmental behaviors of selenium in soil of typical selenosis area. China J. Environ. Sci. 2008, 20, 859–864. [Google Scholar] [CrossRef]
- Fordyce, F.M. Selenium geochemistry and health. Ambio 2007, 36, 94–97. [Google Scholar] [CrossRef]
- Taylor, S.R.; McLennan, S.M. The geochemical evolution of the continental crust. Rev. Geophys. 1995, 33, 241–265. [Google Scholar] [CrossRef]
- Lantzy, R.J.; Mackenzie, F.T. Atmospheric trace metals: Global cycles and assessment of man’s impact. Geochim. Cosmochim. Acta 1979, 43, 511–525. [Google Scholar] [CrossRef]
- Mosher, B.W.; Duce, R.A. A global atmospheric selenium budget. J. Geophys. Res. 1987, 92, 13289–13298. [Google Scholar] [CrossRef]
- Nriagu, J.O.; Pacyna, J.M. Quantitative assessment of worldwide contamination of air, water and soils by trace metals. Nature 1988, 333, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Nriagu, J.O. A global assessment of natural sources of atmospheric trace metals. Nature 1989, 338, 47–49. [Google Scholar] [CrossRef]
- Pacyna, J.M.; Pacyna, E.G. An assessment of global and regional emissions of trace metals to the atmosphere from anthropogenic sources worldwide. Environ. Rev. 2001, 9, 269–298. [Google Scholar] [CrossRef]
- Ross, H.B. An atmospheric selenium budget for the region 30° N to 90° N. Tellus 1985, 37B, 78–90. [Google Scholar] [CrossRef]
- Amouroux, D.; Liss, P.S.; Tessier, E.; Hamren-Larsson, M.; Donard, O.F.X. Role of oceans as biogenic sources of selenium. Earth Planet. Sci. Lett. 2001, 189, 277–283. [Google Scholar] [CrossRef]
- Sievert, S.M.; Kiene, R.P.; Schulz-Vogt, H.N. The sulfur cycle. Oceanography 2007, 20, 117–123. [Google Scholar] [CrossRef]
- Yoch, D.C. Dimethylsulfoniopropionate: Its sources, role in the marine food web, and biological degradation to dimethylsulfide. Appl. Environ. Microbiol. 2002, 68, 5804–5815. [Google Scholar] [CrossRef] [PubMed]
- Vriens, B.; Lenz, M.; Charlet, L.; Berg, M.; Winkel, L.H.E. Natural wetland emissions of methylated trace elements. Nat. Commun. 2014, 5, 3035. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, F.; Lantzy, R.; Paterson, V. Global trace metal cycles and predictions. J. Int. Assoc. Math. Geol. 1979, 11, 99–142. [Google Scholar] [CrossRef]
- Amouroux, D.; Donard, O.F.X. Maritime emission of selenium to the atmosphere in eastern mediterranean seas. Geophys. Res. Lett. 1996, 23, 1777–1780. [Google Scholar] [CrossRef]
- Cutter, G.A. Metalloids in wet deposition on bermuda: concentrations, sources, and fluxes. J. Geophys. Res. 1993, 98, 16777–16786. [Google Scholar] [CrossRef]
- Scudlark, J.R.; Conko, K.M.; Church, T.M. Atmospheric wet deposition of trace elements to Chesapeake Bay: CBAD study year 1 results. Atmos. Environ. 1994, 28, 1487–1498. [Google Scholar] [CrossRef]
- Haygarth, P.M.; Fowler, D.; Stvirup, S.; Davison, B.M.; Jones, K.C. Determination of gaseous and particulate selenium over a rural grassland in the U.K. Atmos. Environ. 1994, 28, 3655–3663. [Google Scholar] [CrossRef]
- Golomb, D.; Ryan, D.; Eby, N.; Underhill, J.; Zemba, S. Atmospheric deposition of toxics onto Massachusetts Bay—I. Metals. Atmos. Environ. 1997, 31, 1349–1359. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, Y.; Yue, T.; Li, Y.; Wai, K.M.; Wang, W. Origin and distribution of trace elements in high-elevation precipitation in southern China. Environ. Sci. Pollut. Res. 2012, 19, 3389–3399. [Google Scholar] [CrossRef] [PubMed]
- Sakata, M.; Marumoto, K.; Narukawa, M.; Asakura, K. Regional variations in wet and dry deposition fluxes of trace elements in Japan. Atmos. Environ. 2006, 40, 521–531. [Google Scholar] [CrossRef]
- Låg, J.; Steinnes, E. Regional distribution of selenium and arsenic in humus layers of Norwegian forest soils. Geoderma 1978, 20, 3–14. [Google Scholar] [CrossRef]
- Blazina, T.; Sun, Y.; Voegelin, A.; Lenz, M.; Berg, M.; Winkel, L.H.E. Terrestrial selenium distribution in China is potentially linked to monsoonal climate. Nat Commun. 2014, 5, 4717. [Google Scholar] [CrossRef] [PubMed]
- Haygarth, P.M.; Cooke, A.I.; Jones, K.C.; Harrison, A.F.; Johnston, A.E. Long-term change in the biogeochemical cycling of atmospheric selenium: Deposition to plants and soil. J. Geophys. Res. 1993, 98, 16769–16776. [Google Scholar] [CrossRef]
- Cao, S.; Duan, X.; Zhao, X.; Ma, J.; Dong, T.; Huang, N.; Sun, C.; He, B.; Wei, F. Health risks from the exposure of children to As, Se, Pb and other heavy metals near the largest coking plant in China. Sci. Total Environ. 2014, 472, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Ranville, M.A.; Cutter, G.A.; Buck, C.S.; Landing, W.M.; Cutter, L.S.; Resing, J.A.; Flegal, A.R. Aeolian contamination of Se and ag in the north pacific from asian fossil fuel combustion. Environ. Sci. Technol. 2010, 44, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Yang, Z.; Lv, Y.; Hou, Q.; Xia, X.; Feng, H.; Zhang, M.; Jin, L.; Kan, Z. The origin and geochemical cycle of soil selenium in a Se-rich area of China. J. Geochem. Explor. 2014, 139, 97–108. [Google Scholar] [CrossRef]
- Gall, H.E.; Sassman, S.A.; Lee, L.S.; Jafvert, C.T. Hormone discharges from a midwest tile-drained agroecosystem receiving animal wastes. Environ. Sci. Technol. 2011, 45, 8755–8764. [Google Scholar] [CrossRef] [PubMed]
- Tarkalson, D.D.; Payero, J.O.; Ensley, S.M.; Shapiro, C.A. Nitrate accumulation and movement under deficit irrigation in soil receiving cattle manure and commercial fertilizer. Agric. Water Manag. 2006, 85, 201–210. [Google Scholar] [CrossRef]
- Van Donk, S.J.; Biswas, S.; Kranz, W.L.; Snow, D.D.; Bartelt-Hunt, S.L.; Mader, T.L.; Shapiro, C.A.; Shelton, D.P.; Tarkalson, D.D.; Zhang, T.C.L.; et al. Transport of steroid hormones in the vadose zone after land application of beef cattle manure. Trans. ASABE 2013, 56, 1327–1338. [Google Scholar]
- Wen, H.; Carignan, J. Selenium isotopes trace the source and redox processes in the black shale-hosted Se-rich deposits in China. Geochim. Cosmochim. Acta 2011, 75, 1411–1427. [Google Scholar] [CrossRef]
- Wen, H.; Carignan, J.; Chu, X.; Fan, H.F.; Cloquet, C.; Huang, J.; Zhang, Y.; Chang, H. Selenium isotopes trace anoxic and ferruginous seawater conditions in the Early Cambrian. Chem. Geol. 2014, 390, 164–172. [Google Scholar] [CrossRef]
- Zhu, J-M.; Johnson, T.M.; Clark, S.K.; Zhu, X-K.; Wang, X-L. Selenium redox cycling during weathering of Se-rich shales: A selenium isotope study. Geochim. Cosmochim. Acta 2014, 126, 228–249. [Google Scholar]
- Pogge von Strandmann, P.A.E.; Coath, C.D.; Catling, D.C.; Poulton, S.W.; Elliott, T. Analysis of mass dependent and mass independent selenium isotope variability in black shales. J. Anal. At. Spectrom. 2014, 29, 1648–1659. [Google Scholar] [CrossRef]
- Stüeken, E.E.; Foriel, J.; Nelson, B.K.; Buick, R.; Catling, D.C. Selenium isotope analysis of organic-rich shales: Advances in sample preparation and isobaric interference correction. J. Anal. At. Spectrom. 2013, 28, 1734–1749. [Google Scholar] [CrossRef]
- Ellis, D.R.; Salt, D.E. Plants, selenium and human health. Curr. Opin. Plant Biol. 2003, 6, 273–279. [Google Scholar] [CrossRef]
- Schilling, K.; Johnson, T.M.; Mason, P.R.D. A sequential extraction technique for mass-balanced stable selenium isotope analysis of soil samples. Chem. Geol. 2014, 381, 125–130. [Google Scholar] [CrossRef]
- Schilling, K.; Wilcke, W. A method to quantitatively trap volatilized organoselenides for stable selenium isotope analysis. J. Environ. Qual. 2011, 40, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Sposito, G. The Chemistry of Soils; Oxford University Press: Oxford, UK, 1989; p. 277. [Google Scholar]
- Le Hécho, I.; Tolu, J.; Thiry, Y.; Bueno, M.; Potin-Gautier, M. Influence of selenium speciation and fractionation on its mobility in soils. In Competitive Sorption and Transport of Heavy Metals in Soils and Geological Media; Magdi Selim, H., Ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 216–230. [Google Scholar]
- Weng, L.; Vega, F.A.; Supriatin, S.; Bussink, W.; Van Riemsdijk, W.H. Speciation of Se and DOC in soil solution and their relation to Se bioavailability. Environ. Sci. Technol. 2011, 45, 262–267. [Google Scholar]
- Elrashidi, M.A.; Adriano, D.C.; Lindsay, W.L. Solubility, speciation, and transformations of selenium in soils. In Agriculture and the Environment; Jacobs, L.W., Ed.; Soil Science Society of America: Madison, WI, USA, 1889; pp. 51–63. [Google Scholar]
- Masscheleyn, P.H.; Delaune, R.D.; Patrick, W.H., Jr. transformations of selenium as affected by sediment oxidation-reduction potential and pH. Environ. Sci. Technol. 1990, 24, 91–96. [Google Scholar] [CrossRef]
- Buchs, B.; Evangelou, M.H.W.; Winkel, L.H.E.; Lenz, M. Colloidal properties of nanoparticular biogenic selenium govern environmental fate and bioremediation effectiveness. Environ. Sci. Technol. 2013, 47, 2401–2407. [Google Scholar] [CrossRef] [PubMed]
- Eszenyi, P.; Sztrik, A.; Babka, B.; Prokisch, J. Elemental, Nano-Sized (100–500 nm) Selenium production by probiotic lactic acid bacteria. Int. J. Biosci. Biochem. Bioinform. 2011, 1, 148–152. [Google Scholar] [CrossRef]
- Dhanjal, S.; Cameotra, S.S. Aerobic biogenesis of selenium nanospheres by Bacillus cereus isolated from coalmine soil. Microb. Cell Fact. 2010, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Tam, K.; Ho, C.T.; Lee, J.H.; Lai, M.; Chang, C.H.; Rheem, Y.; Chen, W.; Hur, H.G.; Myung, N.V. Growth mechanism of amorphous selenium nanoparticles synthesized by Shewanella sp. HN-41. Biosci. Biotechnol. Biochem. 2010, 74, 696–700. [Google Scholar] [CrossRef] [PubMed]
- Lenz, M.; van Aelst, A.C.; Smit, M.; Corvini, P.F.X.; Lens, P.N.L. Biological production of selenium nanoparticles from waste waters. Adv. Mat. Res. 2009, 71–73, 721–724. [Google Scholar] [CrossRef]
- Frost, R.R.; Griffin, R.A. Effect of pH on Adsorption of arsenic and selenium from landfill leachate by clay minerals. Soil Sci. Soc. Am. J. 1977, 41, 53–57. [Google Scholar] [CrossRef]
- Bar-Yosef, B.; Meek, D. Selenium sorption by kaolinite and montmorillonite. Soil Sci. 1987, 144, 11–19. [Google Scholar] [CrossRef]
- Yang, L.; Shahrivari, Z.; Liu, P.K.; Sahimi, M.; Tsotsis, T.T. Removal of trace levels of arsenic and selenium from aqueous solutions by calcined and uncalcined layered double hydroxides (LDH). Ind. Eng. Chem. Res. 2005, 44, 6804–6815. [Google Scholar] [CrossRef]
- Rovira, M.; Giménez, J.; Martínez, M.; Martínez-Lladó, X.; de Pablo, J.; Marti, V.; Duro, L. Sorption of selenium (iv) and selenium (vi) onto natural iron oxides: goethite and hematite. J. Hazard. Mater. 2008, 150, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Eich-Greatorex, S.; Sogn, T.A.; Øgaard, A.F.; Aasen, I. Plant Availability of inorganic and organic selenium fertiliser as influenced by soil organic matter content and pH. Nutr. Cycl. Agroecosyst. 2007, 79, 221–231. [Google Scholar] [CrossRef]
- Hamdy, A.A.; Gissel-Nielsen, G. Fractionation of soil selenium. Zeitschrift für Pflanzenernährung und Bodenkunde 1976, 6, 697–703. [Google Scholar]
- Peak, D.; Sparks, D.L. Mechanisms of selenate adsorption on iron oxides and hydroxides. Environ. Sci. Technol. 2002, 36, 1460–1466. [Google Scholar] [CrossRef] [PubMed]
- Bruggeman, C.; Maes, A.; Vancluysen, J. The interaction of dissolved boom clay and gorleben humic substances with selenium oxyanions (selenite and selenate). Appl. Geochem. 2007, 22, 1371–1379. [Google Scholar] [CrossRef]
- Séby, F.; Gautier, M.P.; Lespes, G.; Astruc, M. Selenium speciation in soils after alkaline extraction. Sci. Total Environ. 1997, 207, 81–90. [Google Scholar] [CrossRef]
- Dhillon, K.S.; Dhillon, S.K. Adsorption–desorption reactions of selenium in some soils of India. Geoderma 1999, 93, 19–31. [Google Scholar] [CrossRef]
- Dhillon, S.K.; Hundal, B.K.; Dhillon, K.S. Bioavailability of selenium to forage crops in a sandy loam soil amended with se-rich plant materials. Chemosphere 2007, 66, 1734–1743. [Google Scholar] [CrossRef] [PubMed]
- Darcheville, O.; Février, L.; Haichar, F.Z.; Berge, O.; Martin-Garin, A.; Renault, P. Aqueous, solid and gaseous partitioning of selenium in an oxic sandy soil under different microbiological states. J. Environ. Radioact. 2008, 99, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Shand, C.A.; Eriksson, J.; Dahlin, A.S.; Lumsdon, D.G. Selenium concentrations in national inventory soils from scotland and sweden and their relationship with geochemical factors. J. Geochem. Explor. 2012, 121, 4–14. [Google Scholar] [CrossRef]
- Levesque, M. Some aspects of selenium relationships in eastern canadian soils and plants. Can. J. Soil Sci. 1974, 54, 205–214. [Google Scholar] [CrossRef]
- Gustafsson, J.P.; Johnsson, L. The association between selenium and humic substances in forested ecosystems—laboratory evidence. Appl. Organomet. Chem. 1994, 8, 141–147. [Google Scholar] [CrossRef]
- Coppin, F.; Chabroullet, C.; Martin-Garin, A. Selenite interactions with some particulate organic and mineral fractions isolated from a natural grassland soil. Eur. J. Soil Sci. 2009, 60, 369–376. [Google Scholar] [CrossRef]
- Jordan, N.; Marmier, N.; Lomenech, C.; Giffaut, E.; Ehrhardt, J.J. Competition between selenium (iv) and silicic acid on the hematite surface. Chemosphere 2009, 75, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Van Dorst, S.H.; Peterson, P.J. Selenium speciation in the soil solution and its relevance to plant uptake. J. Sci. Food Agric. 1984, 35, 601–605. [Google Scholar] [CrossRef]
- Abrams, M.M.; Shennan, C.; Zasoski, R.J.; Burau, R.G. Selenomethionine uptake by wheat seedlings. Agron. J. 1990, 82, 1127–1130. [Google Scholar] [CrossRef]
- Qin, H.B.; Zhu, J.M.; Su, H. Selenium fractions in organic matter from se-rich soils and weathered stone coal in selenosis areas of China. Chemosphere 2012, 86, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Tolu, J.; Thiry, Y.; Bueno, M.; Jolivet, C.; Potin-Gautier, M.; Le Hecho, I. Distribution and speciation of ambient selenium in contrasted soils, from mineral to organic rich. Sci. Total Environ. 2014, 479, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Li, H.F.; Lombi, E.; Stroud, J.L.; McGrath, S.P.; Zhao, F.J. Selenium speciation in soil and rice: Influence of water management and Se fertilization. J. Agric. Food Chem. 2010, 58, 11837–11843. [Google Scholar] [CrossRef] [PubMed]
- Tolu, J.; Le Hécho, I.; Bueno, M.; Thiry, Y.; Potin-Gautier, M. Selenium speciation analysis at trace level in soils. Anal. Chim. Acta 2011, 684, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Stroud, J.L.; McGrath, S.P.; Zhao, F.J. Selenium speciation in soil extracts using LC-ICP-MS. Int. J. Environ. Anal. Chem. 2012, 92, 222–236. [Google Scholar] [CrossRef]
- Winkel, L.H.E.; Johnson, C.A.; Lenz, M.; Grundl, T.; Leupin, O.X.; Amini, M.; Charlet, L. Environmental selenium research: From microscopic processes to global understanding. Environ. Sci. Technol. 2011, 46, 571–579. [Google Scholar] [CrossRef] [PubMed]
- United States Geologic Survey. 2014. Mineral resources on-line spatial data. Available online: Http://mrdata.usgs.gov/ds-801/ (accessed on 27 February 2015).
- Reimann, C.; Birke, M.; Demetriades, A.; Filzmoser, P.; OConnor, P. Chemistry of Europe’s agricultural soils. part a: methodology and interpretation of the GEMAS data set; Bundesanstalt für Geowissenschaften und Rohstoffe: Hannover, Germany, 2014; p. 528. [Google Scholar]
- Soltanpour, P.N. Use of ammonium bicarbonate dtpa soil test to evaluate elemental availability and toxicity 1. Commun. Soil Sci. Plant Anal. 1985, 16, 323–338. [Google Scholar] [CrossRef]
- Barrow, N.J.; Whelan, B.R. Testing a mechanistic model. viii. The Effects of time and temperature of incubation on the sorption and subsequent desorption of selenite and selenate by a soil. J. Soil Sci. 1989, 40, 29–37. [Google Scholar] [CrossRef]
- Chilimba, A.D.C.; Young, S.D.; Black, C.R.; Rogerson, K.B.; Ander, E.L.; Watts, M.J.; Lammel, J.; Broadley, M.R. Maize grain and soil surveys reveal suboptimal dietary selenium intake is widespread in Malawi. Sci. Rep. 2011, 1, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gissel-Nielsen, G. Influence of pH and texture of the soil on plant uptake of added selenium. J. Agric. Food Chem. 1971, 19, 1165–1167. [Google Scholar] [CrossRef] [PubMed]
- Johnsson, L. Selenium uptake by plants as a function of soil type, organic matter content and pH. Plant Soil 1991, 133, 57–64. [Google Scholar] [CrossRef]
- De Temmerman, L.; Waegeneers, N.; Thiry, C.; du Laing, G.; Tack, F.; Ruttens, A. Selenium content of Belgian cultivated soils and its uptake by field crops and vegetables. Sci. Total Environ. 2014, 468, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Geering, H.R.; Cary, E.E.; Jones, L.H.P.; Allaway, W.H. Solubility and redox criteria for the possible forms of selenium in soils. Soil Sci. Soc. Am. J. 1968, 32, 35–40. [Google Scholar] [CrossRef]
- Wang, J.; Li, H.; Li, Y.; Yu, J.; Yang, L.; Feng, F.; Chen, Z. Speciation, distribution, and bioavailability of soil selenium in the Tibetan Plateau Kashin–Beck disease area—a case study in Songpan County, Sichuan Province, China. Biol. Trace Elem. Res. 2013, 156, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Alemi, M.H.; Goldhamer, D.A.; Nielsen, D.R. Modeling selenium transport in steady-state, unsaturated soil columns. J. Environ. Qual. 1991, 20, 89–95. [Google Scholar] [CrossRef]
- Yasin, M.; El-Mehdawi, A.F.; Anwar, A.; Pilon-Smits, E.A.; Faisal, M. Microbial-enhanced selenium and iron biofortification of wheat (triticum aestivum l.)—applications in phytoremediation and biofortification. Int J. Phytoremediat. 2015, 17, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Hengl, T.; Mendes de Jesus, J.; MacMillan, R.A.; Batjes, N.H.; Heuvelink, G.B.M.; Ribeiro, E.; Samuel-Rosa, A.; Kempen, B.; Leenaars, J.G.B.; Walsh, M.G.; Gonzalez, M.R. SoilGrids1 km—global soil information based on automated mapping. PLoS ONE 2014, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, Z.; Duan, B.; Hu, C.; Zhao, X.; Guo, Z. Effect of applied sulphur on the uptake by wheat of selenium applied as selenite. Plant Soil 2015, 386, 35–45. [Google Scholar] [CrossRef]
- Broadley, M.R.; Alcock, J.; Alford, J.; Cartwright, P.; Foot, I.; Fairweather-Tait, S.J.; Hart, D.J.; Hurst, R.; Knott, P.; McGrath, S.P.; et al. Selenium biofortification of high-yielding winter wheat (Triticum aestivum L.) by liquid or granular Se fertilisation. Plant Soil 2010, 332, 5–18. [Google Scholar] [CrossRef]
- Hart, D.J.; Fairweather-Tait, S.J.; Broadley, M.R.; Dickinson, S.J.; Foot, I.; Knott, P.; McGrath, S.P.; Mowat, H.; Norman, K.; Scott, P.R.; et al. Selenium concentration and speciation in biofortified flour and bread: retention of selenium during grain biofortification, processing and production of Se-enriched food. Food Chem. 2011, 126, 1771–1778. [Google Scholar] [CrossRef] [PubMed]
- Renkema, H.; Koopmans, A.; Kersbergen, L.; Kikkert, J.; Hale, B.; Berkelaar, E. the effect of transpiration on selenium uptake and mobility in durum wheat and spring canola. Plant Soil 2012, 354, 239–250. [Google Scholar] [CrossRef]
- Huang, Q.; Yu, Y.; Wang, Q.; Luo, Z.; Jiang, R.; Li, H. Uptake kinetics and translocation of selenite and selenate as affected by iron plaque on root surfaces of rice seedlings. Planta 2014, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C. (Ed.) Atlas of Soil Environmental Background Value in the People’s Republic of China; China Environmental Science Press: Beijing, China, 1994; p. 196.
- Rawlins, B.G.; McGrath, S.P.; Scheib, A.J.; Breward, N.; Cave, M.; Lister, T.R.; Ingham, M.; Gowing, C.; Carter, S. The Advanced Soil Geochemical Atlas of England and Wales; British Geological Survey: Keyworth, UK, 2012; p. 227. [Google Scholar]
- Li, H.F.; McGrath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Zayed, A.; Lytle, C.M.; Terry, N. Accumulation and volatilization of different chemical species of selenium by plants. Planta 1998, 206, 284–292. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Mitani, N.; Yamaji, N.; Shen, R.F.; Ma, J.F. Involvement of silicon influx transporter osnip2;1 in selenite uptake in rice. Plant Physiol. 2010, 153, 1871–1877. [Google Scholar] [CrossRef] [PubMed]
- Hopper, J.; Parker, D. Plant availability of selenite and selenate as influenced by the competing ions phosphate and sulfate. Plant Soil 1999, 210, 199–207. [Google Scholar] [CrossRef]
- Schiavon, M.; Pilon, M.; Malagoli, M.; Pilon-Smits, E.A. Exploring the importance of sulfate transporters and ATP sulphurylases for selenium hyperaccumulation—A comparison of Stanleya pinnata and Brassica juncea (Brassicaceae). Front. Plant Sci. 2015, 6, 1–13. [Google Scholar]
- De Souza, M.P.; Lytle, C.M.; Mulholland, M.M.; Otte, M.L.; Terry, N. Selenium assimilation and volatilization from dimethylselenoniopropionate by Indian mustard. Plant Physiol. 2000, 122, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Kikkert, J.; Berkelaar, E. Plant uptake and translocation of inorganic and organic forms of selenium. Arch. Environ. Contam. Toxicol. 2013, 65, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Fournier, E.; Adam-Guillermin, C.; Potin-Gautier, M.; Pannier, F. Selenate bioaccumulation and toxicity in chlamydomonas reinhardtii: Influence of ambient sulphate ion concentration. Aquat. Toxicol. 2010, 97, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Riedel, G.F.; Sanders, J.G. The influence of ph and media composition on the uptake of inorganic selenium by chlamydomonas reinhardtii. Environ. Toxicol. Chem. 1996, 15, 1577–1583. [Google Scholar] [CrossRef]
- Araie, H.; Sakamoto, K.; Suzuki, I.; Shiraiwa, Y. Characterization of the selenite uptake mechanism in the coccolithophore emiliania huxleyi (haptophyta). Plant Cell Physiol. 2011, 52, 1204–1210. [Google Scholar] [CrossRef] [PubMed]
- Lazard, M.; Blanquet, S.; Fisicaro, P.; Labarraque, G.; Plateau, P. Uptake of selenite by saccharomyces cerevisiae involves the high and low affinity orthophosphate transporters. J. Biol. Chem. 2010, 285, 32029–32037. [Google Scholar] [CrossRef] [PubMed]
- McDermott, J.R.; Rosen, B.P.; Liu, Z. Jen1p: A high affinity selenite transporter in yeast. Mol. Biol. Cell 2010, 21, 3934–3941. [Google Scholar] [CrossRef] [PubMed]
- Obata, T.; Araie, H.; Shiraiwa, Y. Bioconcentration mechanism of selenium by a coccolithophorid, emiliania huxleyi. Plant Cell Physiol. 2004, 45, 1434–1441. [Google Scholar] [CrossRef] [PubMed]
- Nickel, A.; Kottra, G.; Schmidt, G.; Danier, J.; Hofmann, T.; Daniel, H. Characteristics of transport of selenoamino acids by epithelial amino acid transporters. Chem. Biol. Interact. 2009, 177, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Bakke, A.M.; Tashjian, D.H.; Wang, C.F.; Lee, S.H.; Bai, S.C.; Hung, S.S.O. Competition between selenomethionine and methionine absorption in the intestinal tract of green sturgeon (Acipenser medirostris). Aquat. Toxicol. 2010, 96, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Watts, S.D.; Torres-Salazar, D.; Divito, C.B.; Amara, S.G. Cysteine transport through excitatory amino acid transporter 3 (eaat3). PLoS ONE 2014, 9, e109245. [Google Scholar] [CrossRef] [PubMed]
- Riedel, G.; Ferrier, D.; Sanders, J. Uptake of selenium by freshwater phytoplankton. Water Air Soil Pollut. 1991, 57–58, 23–30. [Google Scholar] [CrossRef]
- Huber, R.E.; Criddle, R.S. Comparison of the chemical properties of selenocysteine and selenocystine with their sulfur analogs. Arch. Biochem. Biophys. 1967, 122, 164–173. [Google Scholar] [CrossRef]
- Gojkovic, Ž.; Garbayo, I.; Ariza, J.L.G.; Márová, I.; Vílchez, C. Selenium bioaccumulation and toxicity in cultures of green microalgae. Algal Res. 2015, 7, 106–116. [Google Scholar]
- Mason, R.P. The methylation of metals and metalloids in aquatic systems. In Methylation—From DNA, RNA and Histones to Diseases and Treatment; Dricu, A., Ed.; InTech: Rijeka, Croatia, 2012; pp. 71–301. [Google Scholar]
- Thayer, J.S. Review: Biological methylation of less-studied elements. Appl. Organomet. Chem. 2002, 16, 677–691. [Google Scholar] [CrossRef]
- Araie, H.; Shiraiwa, Y. Selenium utilization strategy by microalgae. Molecules 2009, 14, 4880–4891. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.M.; Arthur, J.R. Selenium, selenoproteins and human health: A review. Public Health Nutr. 2001, 4, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Rahmanto, A.S.; Davies, M.J. Selenium-containing amino acids as direct and indirect antioxidants. IUBMB Life 2012, 64, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Hurst, R.; Collings, R.; Harvey, L.J.; King, M.; Hooper, L.; Bouwman, J.; Gurinovic, M.; Fairweather-Tait, S.J. EURRECA-Estimating selenium requirements for deriving dietary reference values. Crit. Rev. Food Sci. Nutr. 2013, 53, 1077–1096. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, T.C. Selenium biochemistry. Annu. Rev. Biochem. 1990, 59, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Beath, O.A.; Gilbert, C.S.; Eppson, H.F. The use of indicator plants in locating seleniferous soils in the Western United States. I. General. Am. J. Bot. 1939, 26, 257–269. [Google Scholar] [CrossRef]
- Rosenfeld, I.; Beath, O.A. Selenium: Geobotany, Biochemistry, Toxicity, and Nutrition; Academic Press: New York, NY, USA, 1964; p. 411. [Google Scholar]
- Cappa, J.J.; Pilon-Smits, E.A.H. Evolutionary aspects of hyperaccumulation. Planta 2014, 239, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.A.; Shrift, A. Selenium-toxicity and tolerance in higher plants. Biol. Rev. Camb. Philos. 1982, 57, 59–84. [Google Scholar] [CrossRef]
- White, P.J.; Bowen, H.C.; Parmaguru, P.; Fritz, M.; Spracklen, W.P.; Spiby, R.E.; Meacham, M.C.; Mead, A.; Harriman, M.; Trueman, L.J.; et al. Interactions between selenium and sulphur nutrition in Arabidopsis thaliana. J. Exp. Bot. 2004, 55, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gladyshev, V.N. Comparative genomics of trace elements: emerging dynamic view of trace element utilization and function. Chem. Rev. 2009, 109, 4828–4861. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.H.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological functions of beneficial elements. Curr. Opin. Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Vickerman, D.B.; Shannon, M.C.; Bañuelos, G.S.; Grieve, C.M.; Trumble, J.T. Evaluation of Atriplex lines for selenium accumulation, salt tolerance and suitability for a key agricultural insect pest. Environ. Pollut. 2002, 120, 463–473. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Vickerman, D.B.; Trumble, J.T.; Shannon, M.C.; Davis, C.D.; Finley, J.W.; Mayland, H.F. Biotransfer possibilities of selenium from plants used in phytoremediation. Int. J. Phytoremediat. 2002, 4, 315–329. [Google Scholar]
- Hanson, B.; Garifullina, G.F.; Lindblom, S.D.; Wangeline, A.; Ackley, A.; Kramer, K.; Norton, A.P.; Lawrence, C.B.; Pilon Smits, E.A.H. Selenium accumulation protects Brassica juncea from invertebrate herbivory and fungal infection. New Phytol. 2003, 159, 461–469. [Google Scholar] [CrossRef]
- Hanson, B.; Lindblom, S.D.; Loeffler, M.L.; Pilon-Smits, E.A.H. Selenium protects plants from phloem feeding aphids due to both deterrence and toxicity. New Phytol. 2004, 162, 655–662. [Google Scholar] [CrossRef]
- Freeman, J.L.; Lindblom, S.D.; Quinn, C.F.; Fakra, S.; Marcus, M.A.; Pilon-Smits, E.A.H. Selenium accumulation protects plants from herbivory by Orthoptera via toxicity and deterrence. New Phytol. 2007, 175, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Quinn, C.F.; Freeman, J.L.; Reynolds, R.J.B.; Cappa, J.J.; Fakra, S.C.; Marcus, M.A.; Lindblom, S.D.; Quinn, E.K.; Bennett, L.E.; Pilon-Smits, E.A.H. Selenium hyperaccumulation offers protection from cell disruptor herbivores. Plant Physiol. 2010, 153, 1630–1652. [Google Scholar] [CrossRef] [PubMed]
- Quinn, C.F.; Freeman, J.L.; Galeas, M.L.; Klamper, E.M.; Pilon-Smits, E.A.H. The role of selenium in protecting plants against prairie dog herbivory: Implications for the evolution of selenium hyperaccumulation. Oecologia 2008, 155, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.L.; Quinn, C.F.; Lindblom, S.D.; Klamper, E.M.; Pilon-Smits, E.A.H. Selenium protects the hyperaccumulator Stanleya pinnata against black-tailed prairie dog herbivory in native seleniferous habitats. Am. J. Bot. 2009, 96, 1075–1085. [Google Scholar] [CrossRef] [PubMed]
- El Mehdawi, A.F.; Quinn, C.F.; Pilon-Smits, E.A.H. Effects of selenium hyperaccumulation on plant–plant interactions: Evidence for elemental allelopathy. New Phytol. 2011, 191, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Prins, C.N.; Hantzis, L.J.; Quinn, C.F.; Pilon-Smits, E.A.H. Effects of Selenium accumulation on reproductive functions in Brassica juncea and Stanleya pinnata. J. Exp. Bot. 2011, 247. [Google Scholar] [CrossRef] [PubMed]
- Quinn, C.F.; Prins, C.N.; Freeman, J.L.; Gross, A.M.; Hantzis, L.J.; Reynolds, R.J.B.; Yang, S.; Covey, P.A.; Bañuelos, G.S.; Pickering, I.J.; et al. Selenium accumulation in flowers and its effects on pollination. New Phytol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Arvy, M.P. Selenate and selenite uptake and translocation in bean plants (Phaseolus vulgaris). J. Exp. Bot. 1993, 44, 1083–1087. [Google Scholar] [CrossRef]
- Parker, D.R.; Page, A.L.; Bell, P.F. Contrasting selenate-sulfate interactions in selenium-accumulating and nonaccumulating plant species. Soil Sci. Soc. Am. J. 1992, 56, 1818–1824. [Google Scholar] [CrossRef]
- Feist, L.J.; Parker, D.R. Ecotypic variation in selenium accumulation among populations of Stanleya pinnata. New Phytol. 2001, 149, 61–69. [Google Scholar] [CrossRef]
- Harris, J.; Schneberg, K.A.; Pilon-Smits, E.A. Sulfur-selenium-molybdenum interactions distinguish selenium hyperaccumulator Stanleya pinnata from non-hyperaccumulator Brassica juncea (Brassicaceae). Planta 2014, 239, 479–491. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Bowen, H.C.; Marshall, B.; Broadley, M.R. Extraordinarily high leaf selenium to sulfur ratios define ‘Se-accumulator’ plants. Ann. Bot. London 2007, 100, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.L.; Zhang, L.H.; Marcus, M.A.; Fakra, S.; Pilon-Smits, E.A.H. Spatial imaging, speciation and quantification of selenium in the hyperaccumulator plants Astragalus bisulcatus and Stanleya pinnata. Plant Physiol. 2006, 142, 124–134. [Google Scholar] [CrossRef] [PubMed]
- de Souza, M.P.; Pilon-Smits, E.A.H.; Lytle, C.M.; Hwang, S.; Tai, J.C.; Honma, T.S.U.; Yeh, L.; Terry, N. Rate-limiting steps in selenium volatilization by Brassica juncea. Plant Physiol. 1998, 117, 1487–1494. [Google Scholar] [CrossRef] [PubMed]
- Van Hoewyk, D.; Garifullina, G.F.; Ackley, A.R.; Abdel-Ghany, S.E.; Marcus, M.A.; Fakra, S.; Ishiyama, K.; Inoue, E.; Pilon, M.; et al. Overexpression of AtCpNifS enhances selenium tolerance and accumulation in Arabidopsis. Plant Physiol. 2005, 139, 1518–1528. [Google Scholar] [CrossRef] [PubMed]
- Neuhierl, B.; Bock, A. On the mechanism of selenium tolerance in selenium-accumulating plants: Purification and characterization of a specific selenocysteine methyltransferase from cultured cells of Astragalus bisulcatus. Eur. J. Biochem. 1996, 239, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.L.; Quinn, C.F.; Marcus, M.A.; Fakra, S.; Pilon-Smits, E.A.H. Selenium tolerant diamondback moth disarms hyperaccumulator plant defense. Curr. Biol. 2006, 16, 2181–2192. [Google Scholar] [CrossRef] [PubMed]
- Galeas, M.L.; Zhang, L.H.; Freeman, J.L.; Wegner, M.; Pilon-Smits, E.A.H. Seasonal fluctuations of selenium and sulfur accumulation in selenium hyperaccumulators and related non-accumulators. New Phytol. 2007, 173, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Bentley, R.; Chasteen, T.G. Microbial methylation of metalloids: Arsenic, antimony, and bismuth. Microbiol. Mol. Biol. Rev. 2002, 66, 250–271. [Google Scholar] [CrossRef] [PubMed]
- Meija, J.; Montes-Bayón, M.; Le Duc, D.L.; Terry, N.; Caruso, J.A. Simultaneous monitoring of volatile selenium and sulfur species from se accumulating plants (wild type and genetically modified) by gc/ms and gc/icpms using solid-phase microextraction for sample introduction. Anal. Chem. 2002, 74, 5837–5844. [Google Scholar] [CrossRef] [PubMed]
- Dungan, R.S.; Frankenberger, W.T. Biotransformations of selenium by enterobacter cloacae sld1a-1: Formation of dimethylselenide. Biogeochemistry 2001, 55, 73–86. [Google Scholar] [CrossRef]
- Brady, J.M.; Tobin, J.M.; Gadd, G.M. Volatilization of selenite in aqueous medium by a penicillium species. Mycol. Res. 1996, 100, 955–961. [Google Scholar] [CrossRef]
- Neumann, P.M.; de Souza, M.P.; Pickering, I.J.; Terry, N. Rapid microalgal metabolism of selenate to volatile dimethylselenide. Plant Cell Environ. 2003, 26, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Ganther, H.E.; Levander, O.A.; Baumann, C.A. Dietary control of selenium volatilization in the rat. J. Nutr. 1966, 88, 55–60. [Google Scholar] [PubMed]
- Foster, S.J.; Kraus, R.J.; Ganther, H.E. Formation of dimethyl selenide and trimethylselenonium from selenobetaine in the rat. Arch. Biochem. Biophys. 1986, 247, 12–19. [Google Scholar] [CrossRef]
- Kremer, D.; Ilgen, G.; Feldmann, J. Gc-icp-ms determination of dimethylselenide in human breath after ingestion of se-77-enriched selenite: Monitoring of in vivo methylation of selenium. Anal. Bioanal. Chem. 2005, 383, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Alarcon, M.; Cabrera-Vique, C. Selenium in food and the human body: A review. Sci. Total Environ. 2008, 400, 115–141. [Google Scholar] [CrossRef] [PubMed]
- Challenger, F.; Lisle, D.B.; Dransfield, P.B. Studies on biological methylation. Part xiv. The formation of trimethylarsine and dimethyl selenide in mould cultures from methyl sources containing 14C. J. Chem. Soc. (Resumed) 1954, 1954, 1760–1771. [Google Scholar] [CrossRef]
- Doran, J.W.; Alexander, M. Microbial formation of volatile selenium compounds in soil. Soil Sci. Soc. Am. J. 1977, 41, 70–73. [Google Scholar] [CrossRef]
- Reamer, D.C.; Zoller, W.H. Selenium biomethylation products from soil and sewage sludge. Science 1980, 208, 500–502. [Google Scholar] [CrossRef] [PubMed]
- Ansede, J.H.; Yoch, D.C. Comparison of selenium and sulfur volatilization by dimethylsulfoniopropionate lyase (DMSP) in two marine bacteria and estuarine sediments. FEMS Microbiol. Ecol. 1997, 23, 315–324. [Google Scholar] [CrossRef]
- Sunda, W.; Kieber, D.J.; Kiene, R.P.; Huntsman, S. An antioxidant function for dmsp and dms in marine algae. Nature 2002, 418, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Stefels, J. Physiological aspects of the production and conversion of dmsp in marine algae and higher plants. J. Sea Res. 2000, 43, 183–197. [Google Scholar] [CrossRef]
- Kubachka, K.M.; Meija, J.; LeDuc, D.L.; Terry, N.; Caruso, J.A. Selenium volatiles as proxy to the metabolic pathways of selenium in genetically modified Brassica juncea. Environ. Sci. Technol. 2007, 41, 1863–1869. [Google Scholar] [CrossRef] [PubMed]
- Wilber, C.G. Toxicology of selenium: A review. Clin. Toxicol. 1980, 17, 171–230. [Google Scholar] [CrossRef] [PubMed]
- Neuhierl, B.; Thanbichler, M.; Lottspeich, F.; Böck, A. A family of s-methylmethionine-dependent thiol/selenol methyltransferases: Role in selenium tolerance and evolutionary relation. J. Biol. Chem. 1999, 274, 5407–5414. [Google Scholar] [CrossRef] [PubMed]
- Gabel-Jensen, C.; Lunoe, K.; Gammelgaard, B. Formation of methylselenol, dimethylselenide and dimethyldiselenide in in vitro metabolism models determined by headspace GC-MS. Metallomics 2010, 2, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Chau, Y.K.; Wong, P.T.S.; Silverberg, B.A.; Luxon, P.L.; Bengert, G.A. Methylation of selenium in aquatic environments. Science 1976, 192, 1130–1131. [Google Scholar] [CrossRef] [PubMed]
- Cooke, T.D.; Bruland, K.W. Aquatic chemistry of selenium—Evidence of biomethylation. Environ. Sci. Technol. 1987, 21, 1214–1219. [Google Scholar] [CrossRef]
- Pécheyran, C.; Amouroux, D.; Donard, O.F.X. Field determination of volatile selenium species at ultra trace levels in environmental waters by on-line purging, cryofocusing and detection by atomic fluorescence spectroscopy. J. Anal. At. Spectrom. 1998, 13, 615–621. [Google Scholar] [CrossRef]
- Jiang, S.; Robberecht, H.; Adams, F. Identification and determination of alkylselenide compounds in environmental air. Atmos. Environ. 1983, 17, 111–114. [Google Scholar] [CrossRef]
- Calle Guntiñas, M.; Ceulemans, M.; Witte, C.; Łobiński, R.; Adams, F. Evaluation of a purge-and-trap injection system for capillary gas chromatography-microwave induced plasma-atomic emission spectrometry for the determination of volatile selenium compounds in water. Microchim. Acta 1995, 120, 73–82. [Google Scholar] [CrossRef]
- Krupp, E.M.; Grümping, R.; Furchtbar, U.R.; Hirner, A.V. Speciation of metals and metalloids in sediments with ltgc/icp-ms. Anal. Bioanal. Chem. 1996, 354, 546–549. [Google Scholar] [CrossRef] [PubMed]
- Campillo, N.; Aguinaga, N.; Vinas, P.; Lopez-Garcia, I.; Hernandez-Cordoba, M. Gas chromatography with atomic emission detection for dimethylselenide and dimethyldiselenide determination in waters and plant materials using a purge-and-trap preconcentration system. J. Chromatogr. A 2005, 1095, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Tessier, E.; Amouroux, D.; Abril, G.; Lemaire, E.; Donard, O.X. Formation and volatilisation of alkyl-iodidesand -selenides in macrotidal estuaries. Biogeochemistry 2002, 59, 183–206. [Google Scholar] [CrossRef]
- Pécheyran, C.; Lalére, B.; Donard, O.F.X. Volatile metal and metalloid species (Pb, Hg, Se) in a European urban atmosphere (Bordeaux, France). Environ. Sci. Technol. 2000, 34, 27–32. [Google Scholar] [CrossRef]
- Amouroux, D.; Donard, O.F.X. Evasion of selenium to the atmosphere via biomethylation processes in the Gironde estuary, France. Mar. Chem. 1997, 58, 173–188. [Google Scholar] [CrossRef]
- Swearingen, J.W., Jr.; Frankel, D.P.; Fuentes, D.E.; Saavedra, C.P.; Vásquez, C.C.; Chasteen, T.G. Identification of biogenic dimethyl selenodisulfide in the headspace gases above genetically modified escherichia coli. Anal. Biochem. 2006, 348, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Burra, R.; Pradenas, G.A.; Montes, R.A.; Vasquez, C.C.; Chasteen, T.G. Production of dimethyl triselenide and dimethyl diselenenyl sulfide in the headspace of metalloid-resistant bacillus species grown in the presence of selenium oxyanions. Anal. Biochem. 2010, 396, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Vriens, B.; Ammann, A.A.; Hagendorfer, H.; Lenz, M.; Berg, M.; Winkel, L.H.E. Quantification of methylated selenium, sulfur and arsenic in the enviroment. PLoS ONE 2014, 9, e102906. [Google Scholar] [CrossRef] [PubMed]
- Stork, A.; Jury, W.A.; Frankenberger, W.T. Accelerated volatilization rates of selenium from different soils. Biol. Trace Elem. Res. 1999, 69, 217–234. [Google Scholar] [CrossRef] [PubMed]
- Karlson, U.; Frankenberger, W.T. Effects of carbon and trace element addition on alkylselenide production by soil. Soil Sci. Soc. Am. J. 1988, 52, 1640–1644. [Google Scholar] [CrossRef]
- Karlson, U.; Frankenberger, W.T. Accelerated rates of selenium volatilization from California soils. Soil Sci. Soc. Am. J. 1989, 53, 749–753. [Google Scholar] [CrossRef]
- Frankenberger, W.T., Jr.; Karlson, U. Volatilization of selenium from a dewatered seleniferous sediment: A field study. J. Ind. Microbiol. 1995, 14, 226–232. [Google Scholar] [CrossRef]
- Lin, Z.Q.; Cervinka, V.; Pickering, I.J.; Zayed, A.; Terry, N. Managing selenium-contaminated agricultural drainage water by the integrated on-farm drainage management system: Role of selenium volatilization. Water Res. 2002, 36, 3150–3160. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Lin, Z.-Q.; Arroyo, I.; Terry, N. Selenium volatilization in vegetated agricultural drainage sediment from the San Luis drain, central California. Chemosphere 2005, 60, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Terry, N.; Carlson, C.; Raab, T.K.; Zayed, A.M. Rates of selenium volatilization among crop species. J. Environ. Qual. 1992, 21, 341–344. [Google Scholar] [CrossRef]
- Diaz, X.; Johnson, W.P.; Wade, A.O.; Naftz, D.L. Volatile selenium flux from the Great Salt Lake, Utah. Environ. Sci. Technol. 2009, 43, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Huang, Z.-Z. Selenium accumulation and selenium tolerance of salt grass from soils with elevated concentrations of Se and salinity. Ecotoxicol. Environ. Saf. 1991, 22, 267–282. [Google Scholar] [CrossRef]
- Dungan, R.S.; Stork, A.; Frankenberger, W.T. A wind tunnel for measuring selenium volatilization under field-like conditions. J. Environ. Qual. 2000, 29, 460–466. [Google Scholar] [CrossRef]
- Zawislanski, P.T.; Jayaweera, G.R.; Frankenberger, W.T., Jr.; Wu, L. The pond 2 selenium volatilization study: A synthesis of five years of experimental results,1990–1995; A joint report to the Bureau of Reclamation; Earth Sciences Devision: Berkeley, CA, USA, 1996. [Google Scholar]
- Lin, Z.-Q.; Hansen, D.; Zayed, A.; Terry, N. Biological selenium volatilization: Method of measurement under field conditions. J. Environ. Qual. 1999, 28, 309–315. [Google Scholar] [CrossRef]
- Martens, D.A.; Suarez, D.L. Soil methylation-demethylation pathways for metabolism of plant-derived selenoamino acids. In Biogeochemistry of Environmentally Important Trace Elements; Cai, Y.B.O.C., Ed.; ACS Symposium Series; ACS: Washington DC, USA, 2003; Volume 835, pp. 355–369. [Google Scholar]
- Moreno-Jiménez, E.; Clemente, R.; Mestrot, A.; Meharg, A.A. Arsenic and selenium mobilisation from organic matter treated mine spoil with and without inorganic fertilisation. Environ. Pollut. 2013, 173, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Zieve, R.; Peterson, P.J. Factors influencing the volatilization of selenium from soil. Sci. Total Environ. 1981, 19, 277–284. [Google Scholar] [CrossRef]
- Frankenberger, W.T.; Karlson, U. Environmental factors affecting microbial production of dimethylselenide in a selenium contaminated sediment. Soil Sci. Soc. Am. J. 1989, 53, 1435–1442. [Google Scholar] [CrossRef]
- Dhillon, K.S.; Dhillon, S.K.; Dogra, R. Selenium accumulation by forage and grain crops and volatilization from seleniferous soils amended with different organic materials. Chemosphere 2010, 78, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Hansen, D.; Duda, P.J.; Zayed, A.; Terry, N. Selenium removal by constructed wetlands: Role of biological volatilization. Environ. Sci. Technol. 1998, 32, 591–597. [Google Scholar] [CrossRef]
- Gao, S.; Tanji, K.K.; Lin, Z.Q.; Terry, N.; Peters, D.W. Selenium removal and mass balance in a constructed flow-through wetland system. J. Environ. Qual. 2003, 32, 1557–1570. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Moore, J.N. Environmental conditions controlling selenium volatilization from a wetland system. Environ. Sci. Technol. 1997, 31, 511–517. [Google Scholar] [CrossRef]
- Thompson-Eagle, E.T.; Frankenberger, W.T. Volatilization of selenium from agricultural evaporation pond water. J. Environ. Qual. 1990, 19, 125–131. [Google Scholar] [CrossRef]
- Thompson-Eagle, E.T.; Frankenberger, W.T. Protein-mediated selenium biomethylation in evaporation pond water. Environ. Toxicol. Chem. 1990, 9, 1453–1462. [Google Scholar] [CrossRef]
- Calderone, S.J.; Frankenberger, W.T.; Parker, D.R.; Karlson, U. Influence of temperature and organic amendments on the mobilization of selenium in sediments. Soil Biol. Biochem. 1990, 22, 615–620. [Google Scholar] [CrossRef]
- Fan, T.W.M.; Higashi, R.M.; Lane, A.N. Biotransformations of selenium oxyanion by filamentous cyanophyte-dominated mat cultured from agricultural drainage waters. Environ. Sci. Technol. 1998, 32, 3185–3193. [Google Scholar] [CrossRef]
- Frankenberger, W.T.; Karlson, U. Soil management factors affecting volatilization of selenium from dewatered sediments. Geomicrobiol. J. 1994, 12, 265–278. [Google Scholar] [CrossRef]
- Guo, L.; Jury, W.A.; Frankenberger, W.T. Measurement of the henry’s constant of dimethyl selenide as a function of temperature. J. Environ. Qual. 2000, 29, 1715–1717. [Google Scholar] [CrossRef]
- Beale, R.; Johnson, M.; Liss, P.S.; Nightingale, P.D. Air-sea exchange of marine trace gases. In Treatise on Geochemistry, 2nd Ed. ed; Elsevier: Oxford, UK, 2014; pp. 53–92. [Google Scholar]
- Zieve, R.; Peterson, P.J. Sorption of dimethylselenide by soils. Soil Biol. Biochem. 1985, 17, 105–107. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winkel, L.H.E.; Vriens, B.; Jones, G.D.; Schneider, L.S.; Pilon-Smits, E.; Bañuelos, G.S. Selenium Cycling Across Soil-Plant-Atmosphere Interfaces: A Critical Review. Nutrients 2015, 7, 4199-4239. https://doi.org/10.3390/nu7064199
Winkel LHE, Vriens B, Jones GD, Schneider LS, Pilon-Smits E, Bañuelos GS. Selenium Cycling Across Soil-Plant-Atmosphere Interfaces: A Critical Review. Nutrients. 2015; 7(6):4199-4239. https://doi.org/10.3390/nu7064199
Chicago/Turabian StyleWinkel, Lenny H.E., Bas Vriens, Gerrad D. Jones, Leila S. Schneider, Elizabeth Pilon-Smits, and Gary S. Bañuelos. 2015. "Selenium Cycling Across Soil-Plant-Atmosphere Interfaces: A Critical Review" Nutrients 7, no. 6: 4199-4239. https://doi.org/10.3390/nu7064199