2.1. The 3D Laser Triangulation Scanner
The automated plant growth measurements were performed using the commercial 3D laser triangulation scanner PlantEye F300 developed by Phenospex B.V. (Heerlen, the Netherlands) (
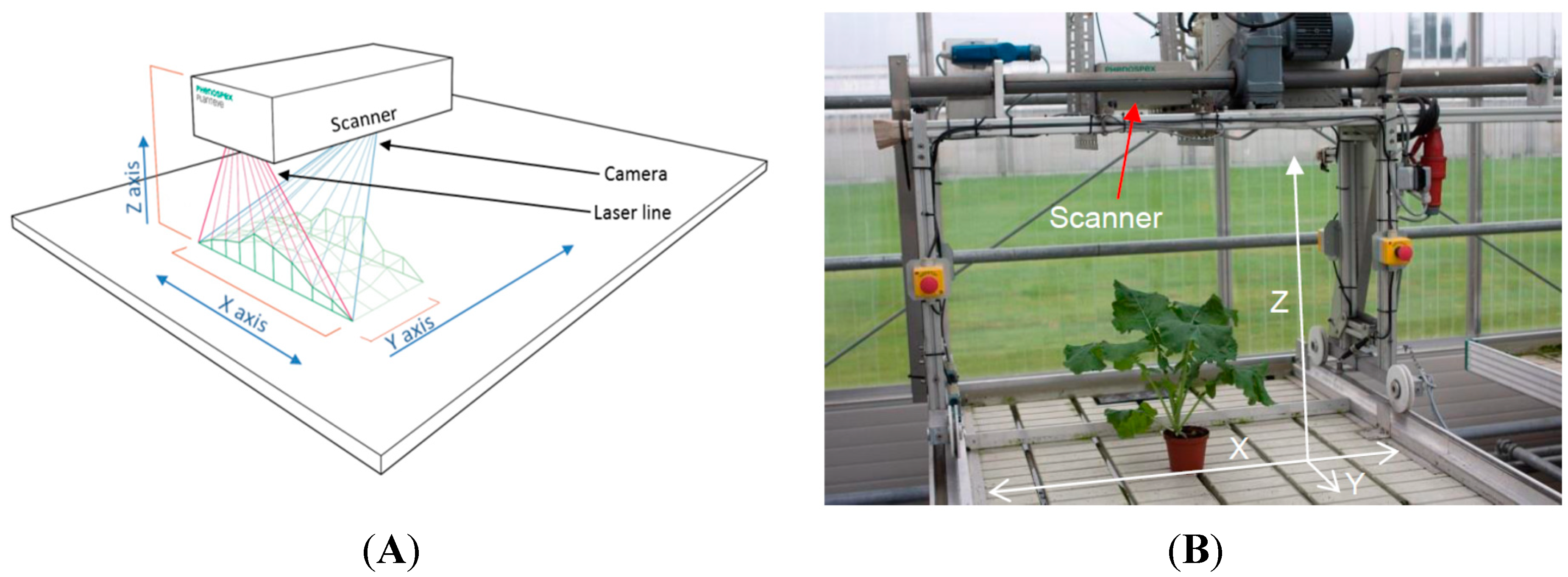
Figure 1A). The sensor projects a laser line in the near infrared (NIR) region of the light spectrum vertically downwards and captures the scattered light with an integrated CMOS-camera.
Figure 1.
(A) Drawing of the 3D scanner, the red lines display the width and projection of the laser line, the blue lines are the projection and width of the camera and the green line/area displays the canopy of a crop stand; (B) The boom system in its starting position with the 3D scanner mounted.
Figure 1.
(A) Drawing of the 3D scanner, the red lines display the width and projection of the laser line, the blue lines are the projection and width of the camera and the green line/area displays the canopy of a crop stand; (B) The boom system in its starting position with the 3D scanner mounted.
A NIR is used to increase data quality since most of the light is reflected from plants. A sunlight filter reduces all artifacts like reflections or background noise from sunlight or other light sources, allowing reproducible measurements under direct sunlight or high irradiances. Moreover, all internal parts like the laser diode and camera is temperature-controlled by a thermoelectric cooler, allowing the operation of the sensor at ambient temperatures of up to 45 °C without cutback or loss of data quality.
The 3D laser triangulation scanner was mounted on a boom (Technical University of Southern Denmark, Odense, Denmark) (
Figure 1B). The boom was placed on a greenhouse table (1.2 × 8 m), with the distance of the scanner to the table of 850 mm, resulting in a scan width (x-direction) of 640 mm. The automated boom can be programmed to run at six different velocities (denoted 1–6), and at specific time points during the day and on specific days, controlled by a digital timer system. To check on the evenness of the velocity of the boom, the time was recorded after every 200 mm at velocity 6. The velocity was measured to be 50.4 mm·s
−1 with a standard error of 1.5 mm·s
−1 over a distance of 6 m. During the scanning process the scanner moves linearly over the plants along the y-axis of the scanning field on the greenhouse table (
Figure 1A, B), and collects images of the projected laser light on the plants.
The resolution in the y-direction depends on the scanning speed of the scanner, which generates 50 xz depth profiles per second, with a resolution of 0.8 mm in the x-direction (width) and a resolution of 0.2 mm in the z-direction (distance from scanner). For instance, if the scanning speed is 50 mm·s
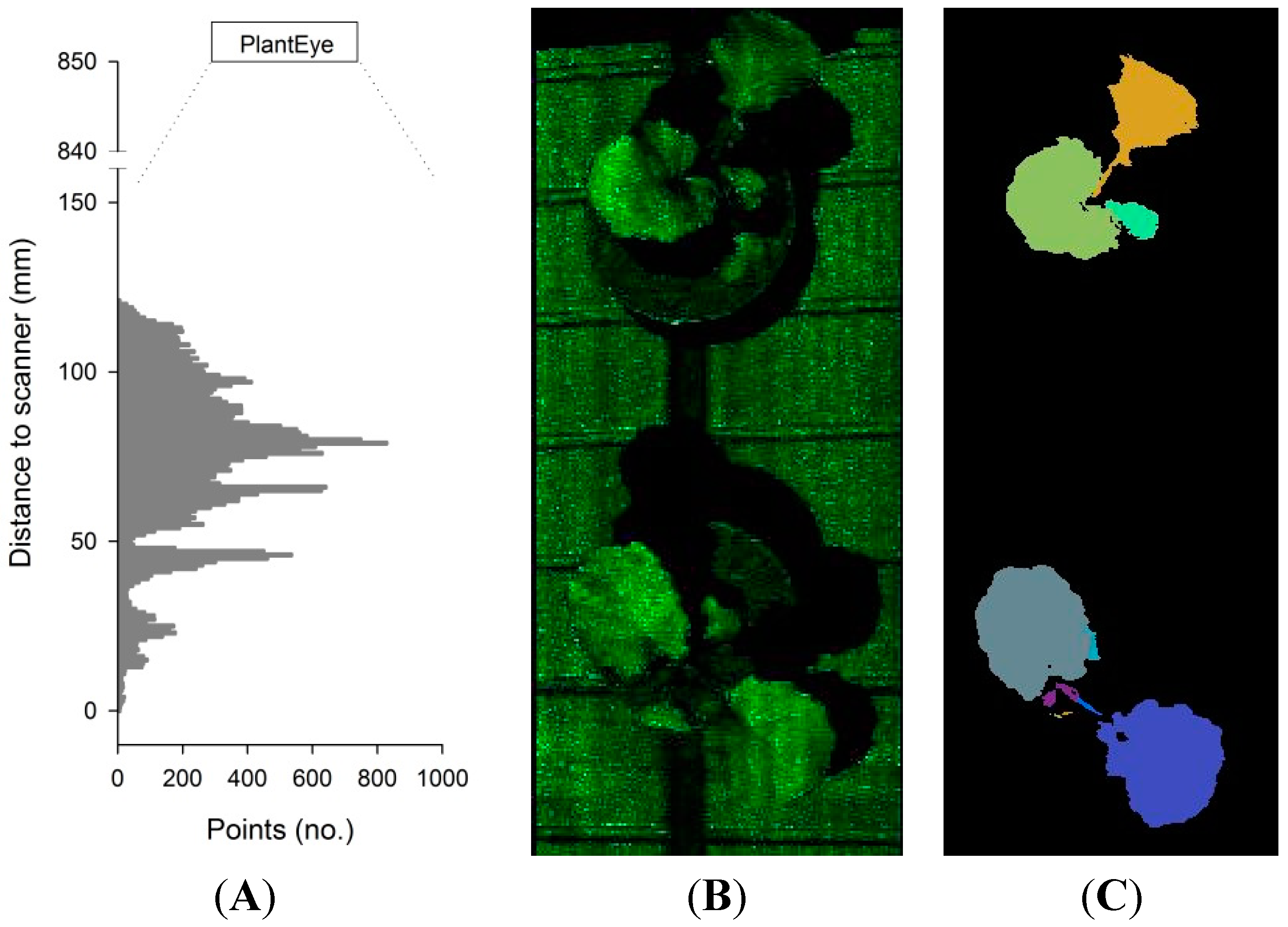
−1, the resolution in the y-direction is 1 mm. The scanning field can be divided into a number of subfields, and from each subfield image, depth profiles of the x-z plane are computed. These depth profiles can be arranged as histograms showing the number of points at a specific distance from the scanner (z-direction) (
Figure 2A), or displayed as a raw 3D point cloud of the subfield canopy (
Figure 2B). From each 3D point cloud, the meshing of neighboring points (segmentation) is automatically carried out (
Figure 2C), and plant height, projected leaf area, 3D leaf area and leaf angle distribution of the subfield canopy are computed.
The height of plants placed in each subfield is calculated from the histograms (
Figure 2A). The points are arranged in percentiles, in relation to their distance from the scanner, and only a part is used for the computation of the plant height. This process is called cropping and is defined in the sensor settings. In most plant species, 80% of the lower points and 10% of the higher points of the histogram are discarded, as the average of the remaining points (between the 80 to 90% percentiles) has been found to give a robust estimate of plant height. The projected leaf area is calculated based on the segmented leaf area in relation to the subfield area (
Figure 2C), whereas the 3D leaf area is computed by considering the distances in 3D taking the steepness of the leaf angles into account. This is a far better estimation of leaf area compared to the projected leaf area which is delivered by 2D imaging.
Figure 2.
(A) Histogram showing the number of points at different distances to the scanner; (B) the raw 3D point cloud of two rapeseed plants and (C) the segmented 3D point cloud.
Figure 2.
(A) Histogram showing the number of points at different distances to the scanner; (B) the raw 3D point cloud of two rapeseed plants and (C) the segmented 3D point cloud.
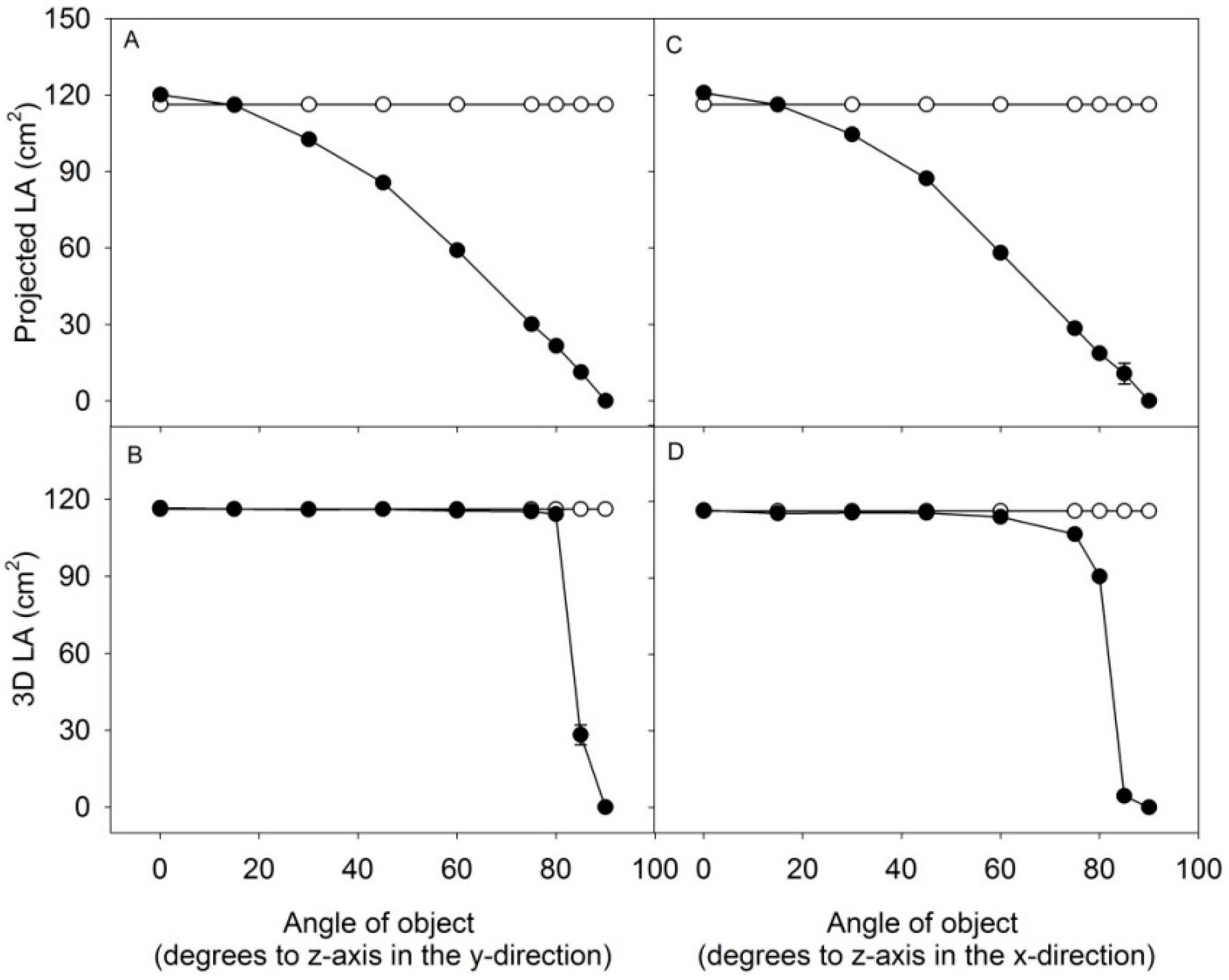
A comparison between the estimate of 3D leaf area and projected leaf area (2D) was made by scanning a flat object with an area of 116.2 cm
2 with the angle of the object towards the scanner increased incrementally in the y-direction (
Figure 3A,B) or x-direction (
Figure 3C,D). The surface of the object was estimated to 116.4 ± 2 cm
2 at an angle of 0° for 3D area and to 118.9 ± 1 cm
2 for 2D area. At an angle of 80° in the y-direction, the 3D area was 107.6 ± 3 cm
2 and the projected area was 22.8 ± 1 cm
2; a similar result was obtained when tilting the object in the x-direction. Above the angle of 80°, the 3D leaf area was incorrectly estimated, especially when the object was tilted in the x-direction. This was caused by insufficient scattering of light back into the camera. The small test demonstrate the advantages of 3D measurements, being sufficiently precise in predicting the area, even at increasing leaf-angles in both the x- and y-direction, compared to 2D which only allows the measurement of the projected leaf area.
Figure 3.
An object with defined area of 116.2 cm2 was tilted stepwise to the z-plane in the y- and x-direction and measured by the 3D scanner. Projected LA and 3D LA of the object was computed at different angles. Area of object (white dots), computed area of object (black dots). Values are average of three ± SE, with SE’s below 1 cm2 for most values.
Figure 3.
An object with defined area of 116.2 cm2 was tilted stepwise to the z-plane in the y- and x-direction and measured by the 3D scanner. Projected LA and 3D LA of the object was computed at different angles. Area of object (white dots), computed area of object (black dots). Values are average of three ± SE, with SE’s below 1 cm2 for most values.
2.2. Experiment 1: Testing a Potential Influence of the Projected Laser Line on Photosynthetic Activity
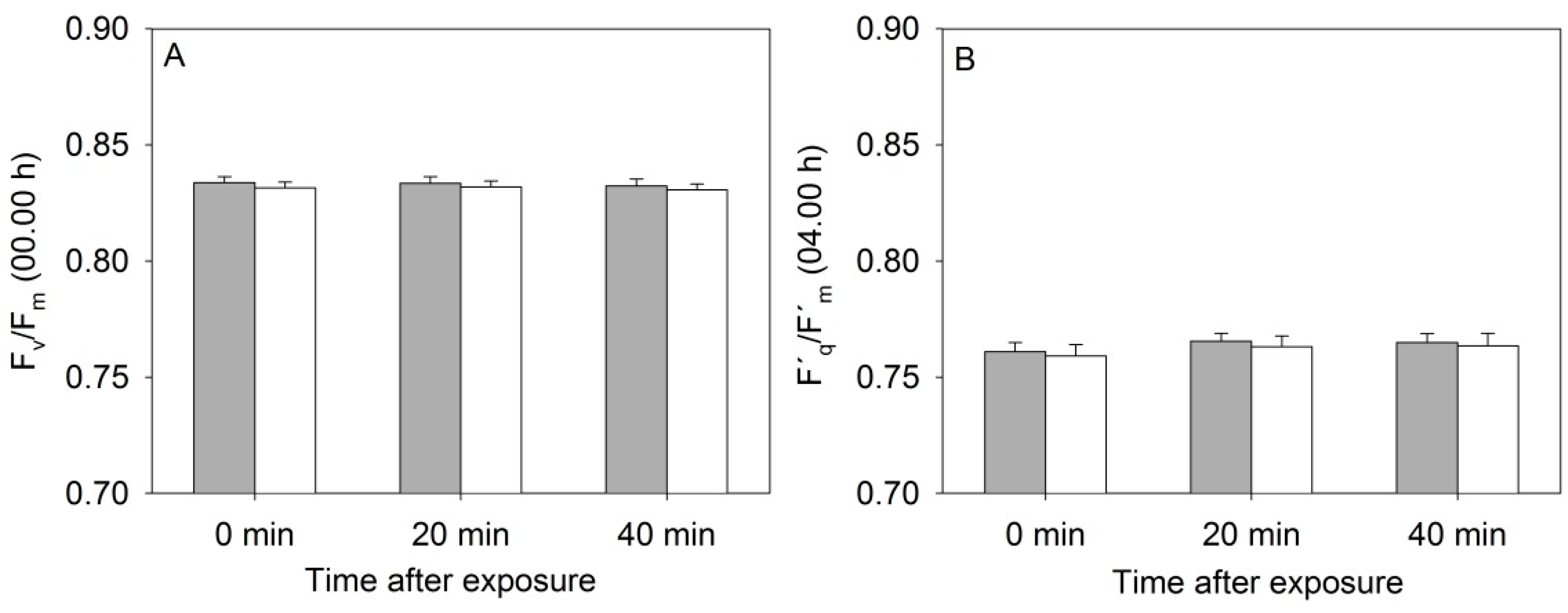
The 3D laser triangulation scanner is equipped with a NIR laser belonging to the laser class 1 M (940 nm), meaning that it is eye-safe to use in all conditions except when passed through magnifying optics.
In order to test whether a laser line of this power had any effects on the plant physiological performance, the PSII photochemistry of rapeseed plants was monitored continuously with a PAM fluorimeter (MONI-PAM, Walz, Effeltrich, Germany) for plants placed underneath the scanner. Half of the laser line was covered using black tape (TESA 4613, tesa A/S, Amsterdam, The Netherlands) and six plants were placed on the table so that three plants were exposed to the laser and three plants were not exposed to the laser. The system was set to measure every 30 min (00:00 h and 00:30 h) at a scanning speed of 20 mm·s
−1. The measurements took 5 min from start to end, and the MONI-PAM was set to measure every 20 min (00:05, 00:25 and 00:45 h). The chlorophyll fluorescence measuring system consisted of six emitter-detector units (MONI-heads), each representing independent fluorimeters. Three MONI-heads were placed directly underneath the path of the laser line, and the three other MONI-heads were placed outside that path. Rapeseed leaves of similar age were fixed in the MONI-heads leaf clips, and measurements of photosynthetic active irradiation in the range of 400–700 nm (PAR, µmol·m
−2·s
−1) using the integrated quantum sensor, maximum photochemical efficiency of PSII during the dark period; F
v/F
m = (F
m − F
0)/F
m, Quantum yield of PSII (Φ
PSII); F´
q/F´
m = (F´
m − F´)/F´
m and the electron transport rate (ETR) during the light period [
14], were recorded continuously as described above. The intensity of the light saturating pulse was 1800 µmol·m
−2·s
−1 and the duration of the pulse was 0.8 s. The measurements were done on eight consecutive days. Every day at 10:00 h the MONI-heads were randomized, and a new part of the leaf was measured, and after four days the black tape was moved to cover the other 50% of the laser line in order to obtain a completely randomized design.
2.3. Experiment 2: Predicting Growth Parameters of Rapeseed by 3D Laser Triangulation
The experiment was conducted to develop a model from which it would be possible to predict destructive growth parameters of rapeseed by using the parameters obtained by 3D laser triangulation. The experiment on rapeseed was carried out from 24 September to 6 October 2013 (plant batch 1), and repeated from 8–29 November 2013 (plant batch 2) in a greenhouse located at the Department of Food Science, University of Aarhus (Aarslev, Denmark, Lat. 55°N). The light period was 20 h per day, supplemented by artificial light provided by high-pressure sodium lamps (SON-T agro, 600 W, Phillips, Eindhoven, The Netherlands) at 130 µmol·m−2·s−1 photosynthetic flux density. The temperature set point was 20 °C with opening of the vents at 25 °C. The plants were watered by flooding for a short period every second day.
Seeds of the rapeseed genotype “DH5” were sown in 11 cm pots containing peat and placed immediately on the table where the 3D scanner was mounted. The field of the 3D scanner was divided into ten longitudinal subfields (640 × 600 mm) with six pots in each subfield, distributed in two rows with three pots in each. The distance between the two rows was 80 mm and the distance between pots in the rows was 60 mm. Plants from each of ten subfields were harvested at growth stages defined by the number of leaves, to allow comparison of destructive and non-destructive measurements at different stages of plant development. The scanning measurements of each subfield were conducted with a scan velocity of the boom of 20 mm·s−1, giving a resolution of less than 1 mm in the scanning direction (y). Scans were carried out every hour over a period of three weeks.
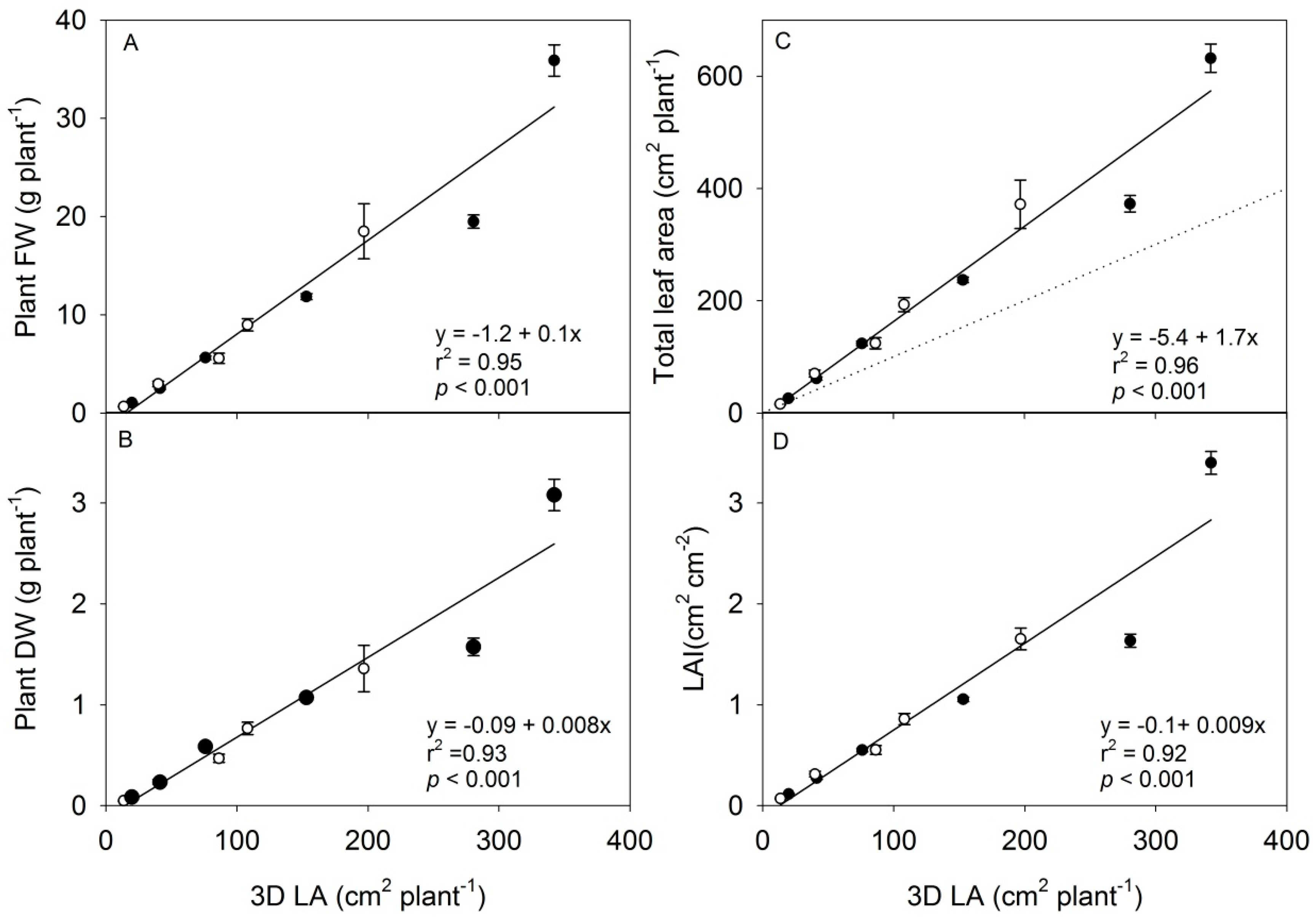
The destructive harvests were carried out at 12:00 h, and all six plants from a subfield were harvested. For each plant, leaves were separated from the petioles, the number of leaves was counted (LN) and the total leaf area (LA) was determined using a leaf area meter (LI-3000, LI-COR, Lincoln, NE). Plant fresh weight (FW) was determined, plant material was dried at 70 °C for 24 h and plant dry weight (DW) was determined. The leaf area index (LAI) was calculated as total leaf area of the six plants per ground area of the subfield (1350 cm2). The average values per plant of the destructive growth measurements (FW, DW, LN, LA and LAI) were related to the estimated values of projected leaf area (cm2·plant−1), 3D leaf area (cm2·plant−1) and height (cm). The estimated values were calculated from the scanning measurements conducted in the middle of the dark period (00:00 h) the night before the destructive harvests. This time point was chosen based on earlier observations that variation in environmental conditions have least effect on the plant structure at this time of day, making the day-to-day measurements of growth most reliable. The calculated parameters were based on four out of the six measured plants to avoid two plants at the edge, which were expanding their leaves outside the plot. These two plants were manually removed from the 3D point cloud. The correlation analysis was carried out on plants with leaf numbers ranging from two to seven (including cotyledons) from six subfields out of ten from each batch (12 in total). In the last four subfields of both batches, the plants had too many overlapping leaves, or leaves were extending out of the plot.
2.4. Experiment 3: Validating the Model on Ten Rapeseed Genotypes
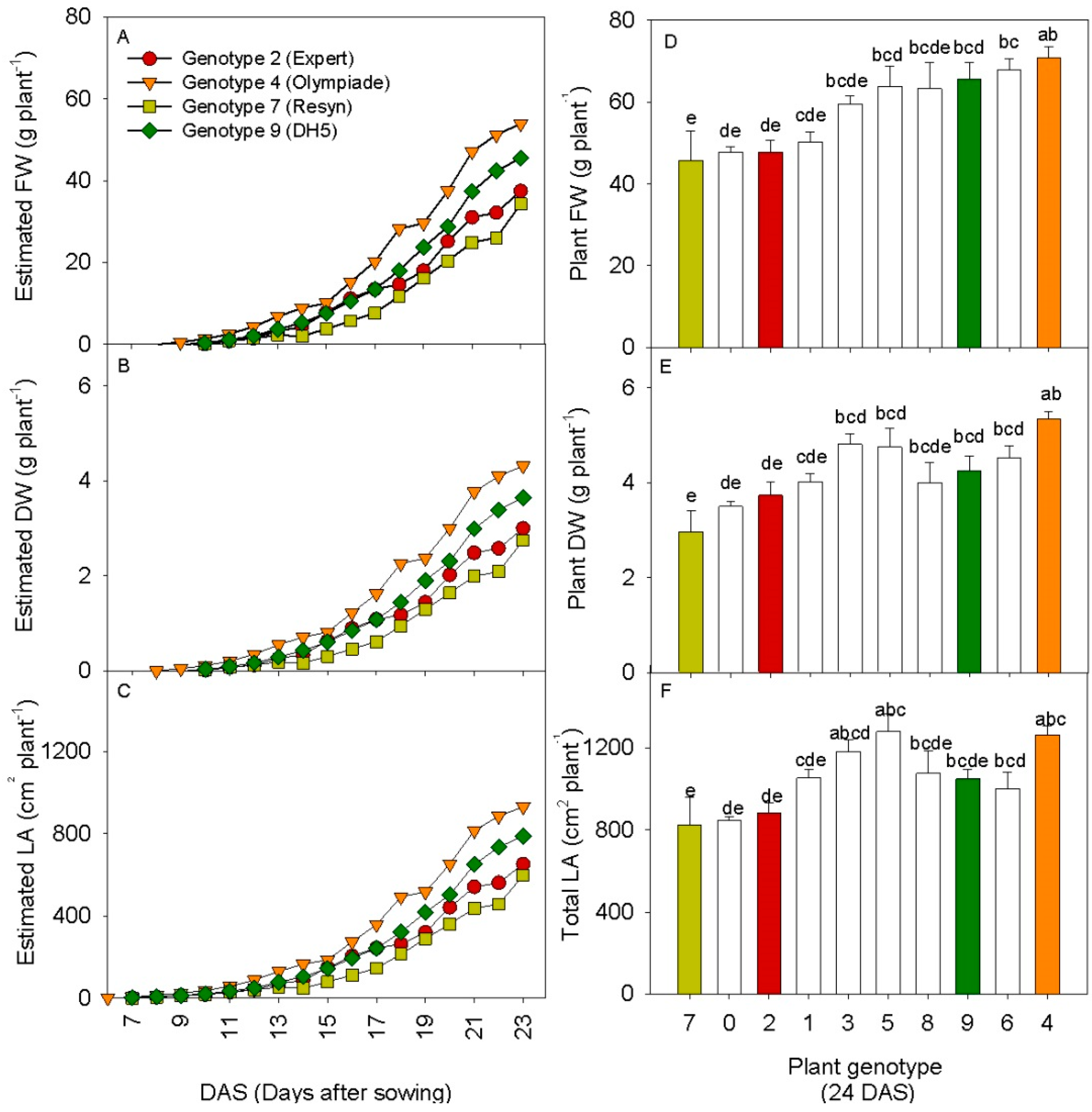
A screening experiment with ten rapeseed genotypes was carried out in order to validate the linear models on the rapeseed genotype “DH5” from experiment 2. The experiment was carried out from 26 July to 19 August 2013 with greenhouse climate set points similar to those for Experiment 2. The seeds of ten rapeseed genotypes, 0. Chuosenshu, 1. Cobra, 2. Expert OSR, 3. Palu, 4. Olympiade, 5. Major, 6. S13, 7. Resyn HO48, 8. Markus and 9. DH5, were sown in 11 cm pots containing peat and placed directly in ten subfields on the table where the scanner was mounted. The distance between the pots was similar to that in experiment 2. Scanning measurements of each subfield were conducted every hour at a scan velocity of 20 mm·s−2, as in experiment 2. At the final harvest 24 days after sowing (DAS), all plants from each subfield were harvested. For each plant, the leaves were counted (LN) and the total leaf area (LA) was determined using a leaf area meter (LI-3000, LI-COR, Lincoln, NE, USA). Plant fresh weight (FW) was determined, plant material was dried at 70 °C for 24 h and plant dry weight (DW) was determined. The linear models obtained in experiment 2 were used to calculate daily values for plant FW, DW and total leaf area, to generate growth curves for the 23 DAS until the 18 August, the day before harvest.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


