Biohydrogen Production from Food Waste: Influence of the Inoculum-To-Substrate Ratio

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrate and Inoculum
2.2. Experimental Set-Up
2.3. Analytical Methods
2.4. Calculations
2.5. Kinetic Model
2.6. Statistical Analysis
3. Results
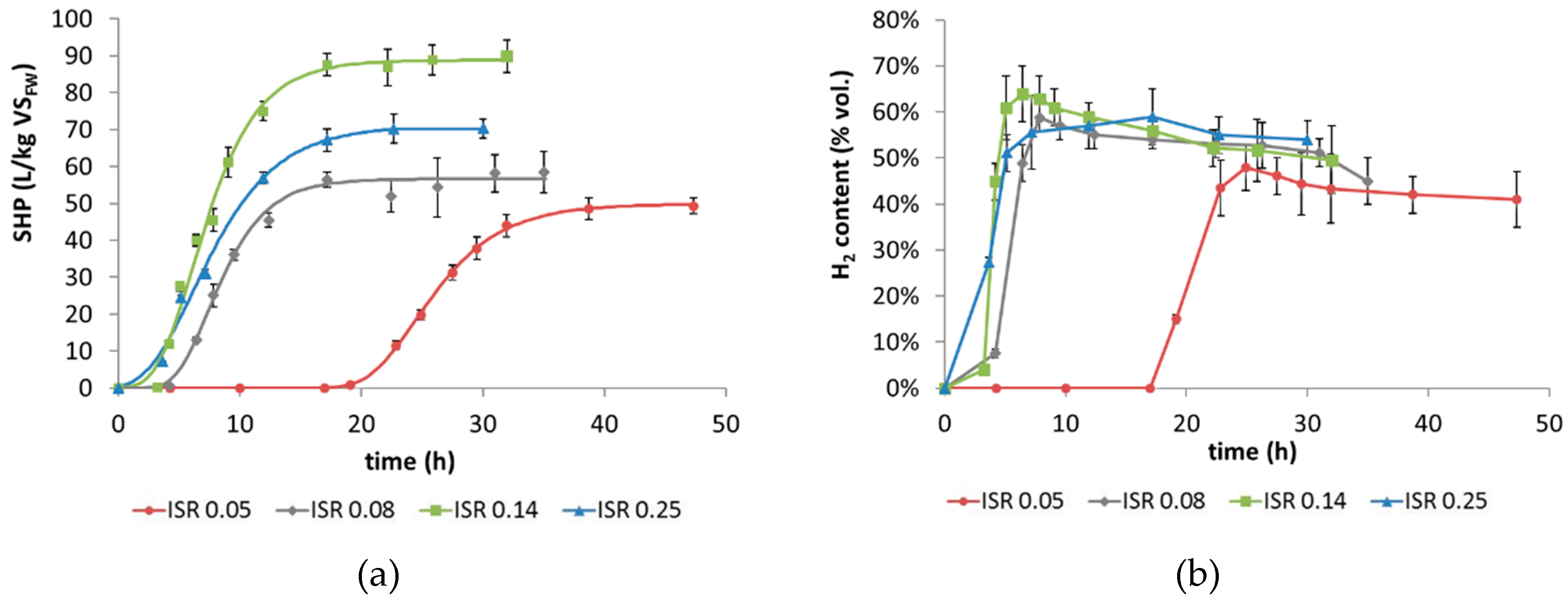
3.1. Hydrogen Production
3.2. Hydrogen Production Kinetics
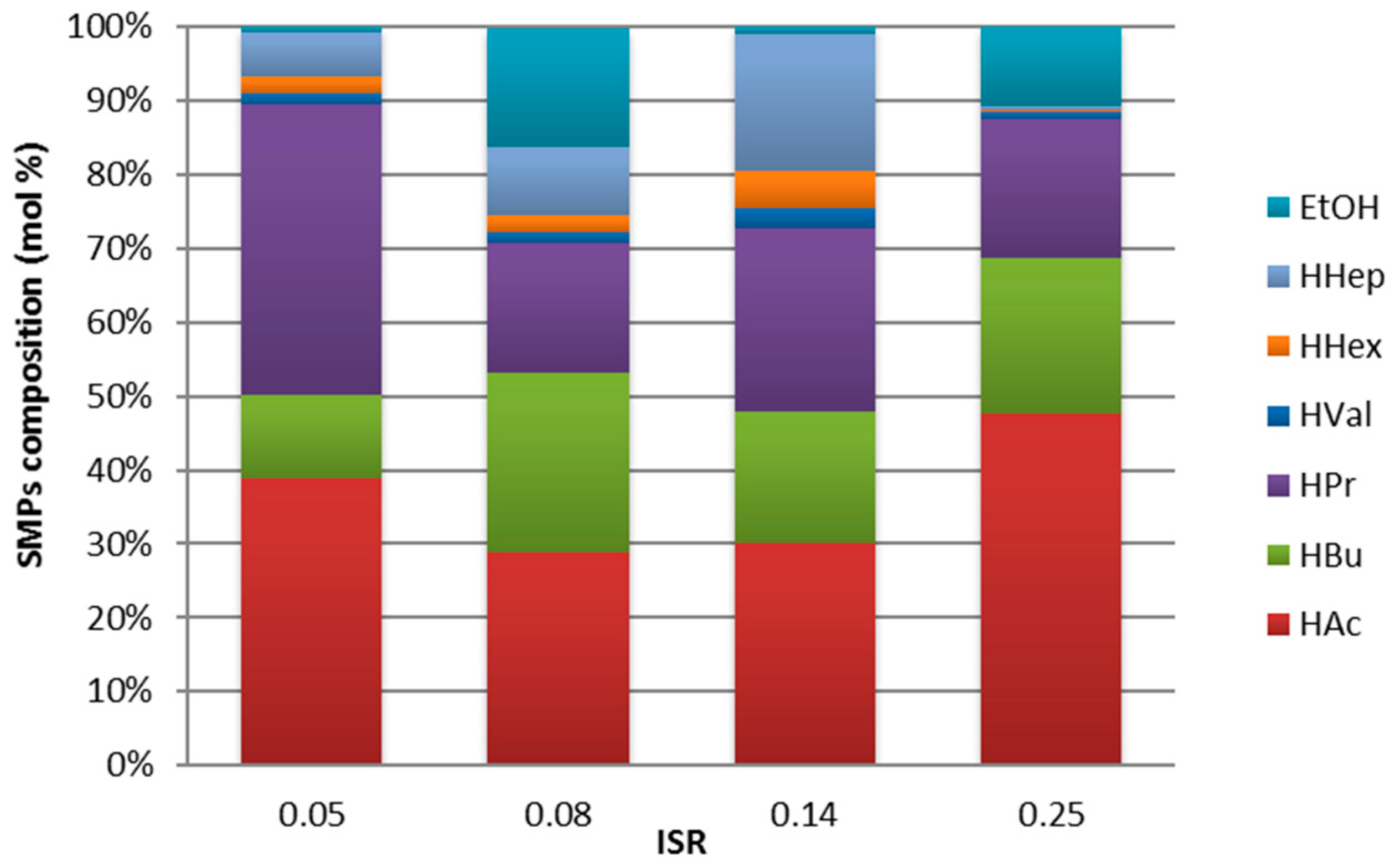
3.3. Fermentation Products and Substrate Conversion
4. Conclusions
- The ISR exerted a remarkable influence on both the process kinetics and the final H2 production yield.
- An appropriate ISR proved to enhance the effects of an optimal operating pH, confirming that fermentative H2 production is a process that requires substrate-specific optimisation of a plurality of operating parameters.
- An ISR of 0.14 proved to be the optimal value for fermentative H2 production from FW, as suggested by the observed performance in terms of SHPmax (88.8 L H2/kg VSFW) and Rmax (10.8 L H2/kg VSFW∙h).
- The main metabolic products included acetate, butyrate, propionate, and ethanol. Several overlapping and competing fermentation pathways likely governed the process, reducing the observed H2 production.
- The high correspondence between OBSH2 and THEOH2 for ISR 0.14 suggests that in this test, the H2 production mostly derived from the typical acetate/butyrate-producing Clostridial fermentation, with the net of the H2 consumption related to propionic fermentation.
- Optimising the ISR provided useful information to support the perspectives for real-scale implementation of fermentative hydrogen production. Among the aspects that would provide the most benefit, the standardisation of tests to estimate the hydrogen production potential from different substrates is of particular importance. The recently published German guideline VDI 4630 (2016) has emphasised the role of the biochemical methane potential test as a reliable approach for the determination of the methane production potential [17,45]; similarly, a biochemical hydrogen potential test could be worth developing as a valuable, simple, and low cost tool to assess the potential, adequacy, and viability of the fermentative hydrogen production process [10,20,46,47].
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Han, S.-K.; Shin, H.-S. Performance of an innovative two-stage process converting food waste to hydrogen and methane. J. Air Waste Manag. Assoc. 2004, 54, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.B.; Pitt, L.; Love, M. Biohydrogen production: Prospects and limitations to practical application. Int. J. Hydrog. Energy 2004, 29, 173–185. [Google Scholar] [CrossRef]
- Antonopoulou, G.; Gavala, H.N.; Skiadas, I.V.; Angelopoulos, K.; Lyberatos, G. Biofuels generation from sweet sorghum: Fermentative hydrogen production and anaerobic digestion of the remaining biomass. Bioresour. Technol. 2008, 99, 110–119. [Google Scholar] [CrossRef] [PubMed]
- De Gioannis, G.; Muntoni, A.; Polettini, A.; Pomi, R.; Spiga, D. Energy recovery from one- and two-stage anaerobic digestion of food waste. Waste Manag. 2017, 68, 595–602. [Google Scholar] [CrossRef] [PubMed]
- De Gioannis, G.; Muntoni, A.; Polettini, A.; Pomi, R. A review of dark fermentative hydrogen production from biodegradable municipal waste fractions. Waste Manag. 2013, 33, 1345–1361. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; He, Q.; Zhao, X.; Wu, D.; Wang, X.; Peng, X. Anaerobic digestion of food waste: Correlation of kinetic parameters with operational conditions and process performance. Biochem. Eng. J. 2018, 130, 1–9. [Google Scholar] [CrossRef]
- Liu, Y. Bioenergenic interpretation on So/Xo ratio in substrate-sufficient batch culture. Water Res. 1996, 30, 2766–2770. [Google Scholar] [CrossRef]
- Stanislaus, M.S.; Zhang, N.; Yuan, Y.; Zheng, H.; Zhao, C.; Hu, X.; Zhu, Q.; Yang, Y. Improvement of biohydrogen production by optimization of pretreatment method and substrate to inoculum ratio from microalgal biomass and digested sludge. Renew. Energy 2018, 127, 670–677. [Google Scholar] [CrossRef]
- Argun, H.; Dao, S. Bio-hydrogen production from waste peach pulp by dark fermentation: Effect of inoculum addition. Int. J. Hydrog. Energy 2017, 42, 2569–2574. [Google Scholar] [CrossRef]
- Ghimire, A.; Sposito, F.; Frunzo, L.; Trably, E.; Escudié, R.; Pirozzi, F.; Lens, P.N.L.; Esposito, G. Effects of operational parameters on dark fermentative hydrogen production from biodegradable complex waste biomass. Waste Manag. 2016, 50, 55–64. [Google Scholar] [CrossRef]
- Cappai, G.; De Gioannis, G.; Muntoni, A.; Polettini, A.; Pomi, R.; Spiga, D. Effect of inoculum to substrate ratio (ISR) on hydrogen production through dark fermentation of food waste. In Proceedings of the Atti del “Sardinia 2015 15th International Waste Management and Landfill Symposium”, Cagliari, Italy, 5–9 October 2015. [Google Scholar]
- De Gioannis, G.; Friargiu, M.; Gioia, M.; Muntoni, A.; Polettini, A.; Pomi, R. Effect of inoculum to substrate ratio (ISR) and pH on fermentative hydrogen production from cheese whey. In Proceedings of the Atti del “Sardinia 2015 15th International Waste Management and Landfill Symposium", Cagliari, Italy, 5–9 October 2015. [Google Scholar]
- Nathao, C.; Sirisukpoka, U.; Pisutpaisal, N. Production of hydrogen and methane by one and two stage fermentation of food waste. Int. J. Hydrog. Energy 2013, 38, 15764–15769. [Google Scholar] [CrossRef]
- Chen, W.; Chen, S.; Kumar, S.; Sung, S. Kinetic study of biological hydrogen production by anaerobic fermentation. Int. J. Hydrog. Energy 2006, 31, 2170–2178. [Google Scholar] [CrossRef]
- Sangyoka, S.; Reungsang, A.; Lin, C.-Y. Optimization of biohydrogen production from sugarcane bagasse by mixed cultures using a statistical method. Sustain. Environ. Res. 2016, 26, 235–242. [Google Scholar] [CrossRef]
- Costa, J.C.; Oliveira, J.V.; Alves, M.M. Response surface design to study the influence of inoculum, particle size and inoculum to substrate ratio on the methane production from Ulex sp. Renew. Energy 2016, 96, 1071–1077. [Google Scholar] [CrossRef] [Green Version]
- Raposo, F.; De la Rubia, M.A.; Fernandez-Cegrì, V.; Borja, R. Biochemical methane potential (BMP) of solid organic substrates: Evaluation of anaerobic biodegradability using data from an international interlaboratory study. Renew. Sustain. Energy Rev. 2011, 16, 861–877. [Google Scholar] [CrossRef]
- Boulanger, A.; Pinet, E.; Bouix, M.; Bouchez, T.; Mansour, A.A. Effect of inoculum to substrate ratio (I/S) on municipal solid waste anaerobic degradation kinetics and potential. Waste Manag. 2012, 32, 2258–2265. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Zhang, R.; El-mashad, H.M.; Sun, H.; Ying, Y. Effect of food to microorganism ratio on biohydrogen production from food waste via anaerobic fermentation. Int. J. Hydrog. Energy 2008, 33, 6968–6975. [Google Scholar] [CrossRef]
- Alibardi, L.; Cossu, R. Composition variability of the organic fraction of municipal solid waste and effects on hydrogen and methane production potentials. Waste Manag. 2015, 36, 147–155. [Google Scholar] [CrossRef]
- Cappai, G.; De Gioannis, G.; Friargiu, M.; Massi, E.; Muntoni, A.; Polettini, A.; Pomi, R.; Spiga, D. An experimental study on fermentative H2 production from food waste as affected by pH. Waste Manag. 2014, 34, 1510–1519. [Google Scholar] [CrossRef]
- American Public Health Association and American Water Works Association. Standard Methods for the Examination of Water and Wastewater, 21th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Voelklein, M.A.; Jacob, A.; O’ Shea, R.; Murphy, J.D. Assessment of increasing loading rate on two-stage digestion of food waste. Bioresour. Technol. 2016, 202, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Fang, H.H.P. Fermentative hydrogen production from wastewater and solid wastes by mixed cultures. Crit. Rev. Environ. Sci. Technol. 2007, 37, 1–39. [Google Scholar] [CrossRef]
- Jungermann, K.; Thauer, R.K.; Leimenstoll, G.; Decker, K. Function of reduced pyridine nucleotide-ferredoxin oxidoreductases in saccharolytic Clostridia. Biochim. Biophys. Acta–Bioenerg. 1973, 305, 268–280. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; van ’t Riet, K. Modeling of the bacterial growth curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [PubMed]
- Lay, J.-J.; Lee, Y.-J.; Noike, T. Feasibility of biological hydrogen production from organic fraction of municipal solid waste. Water Res. 1999, 33, 2579–2586. [Google Scholar] [CrossRef]
- Dong, L.; Zhenhong, Y.; Yongming, S.; Longlong, M. Anaerobic fermentative co-production of hydrogen and methane from an organic fraction of municipal solid waste. Energy Sources Part A Recover. Util. Environ. Eff. 2011, 33, 575–585. [Google Scholar] [CrossRef]
- Siriwongrungson, V.; Zeng, R.J.; Angelidaki, I. Homoacetogenesis as the alternative pathway for H2 sink during thermophilic anaerobic degradation of butyrate under suppressed methanogenesis. Water Res. 2007, 41, 4204–4210. [Google Scholar] [CrossRef]
- Saady, N.M.C. Homoacetogenesis during hydrogen production by mixed cultures dark fermentation: Unresolved challenge. Int. J. Hydrog. Energy 2013, 38, 13172–13191. [Google Scholar] [CrossRef]
- Zhang, C.; Su, H.; Baeyens, J.; Tan, T. Reviewing the anaerobic digestion of food waste for biogas production. Renew. Sustain. Energy Rev. 2014, 38, 383–392. [Google Scholar] [CrossRef]
- Akhlaghi, M.; Boni, M.R.; De Gioannis, G.; Muntoni, A.; Polettini, A.; Rossi, A.; Spiga, D. A parametric response surface study of fermentative hydrogen production from cheese whey. Bioresour. Technol. 2017, 244, 473–483. [Google Scholar] [CrossRef]
- Bundhoo, M.A.Z.; Mohee, R. Inhibition of dark fermentative bio-hydrogen production: A review. Int. J. Hydrog. Energy 2016, 41, 6713–6733. [Google Scholar] [CrossRef]
- Park, W.S.; Hwang, M.H.; Hyun, S.H.; Kim, I.S. Suppression of hydrogen consuming bacteria in anaerobic hydrogen fermentation. Environ. Eng. Res. 2005, 10, 181–190. [Google Scholar] [CrossRef]
- Gottschalk, J.C.; Morris, J.G. The induction of acetone and butanol production in cultures Clostridium acetobutylicum by elevated concentrations of acetate and butyrate. FEMS Microb. Lett. 1981, 12, 385–389. [Google Scholar] [CrossRef]
- Terracciano, J.S.; Kashket, E.R. Intracellular conditions required for initiation of solvent production by Clostridium acetobutylicum. Appl. Environ. Microbiol. 1986, 52, 86–91. [Google Scholar] [PubMed]
- Argun, H.; Kargi, F.; Kapdan, I.K.; Oztekin, R. Batch dark fermentation of powdered wheat starch to hydrogen gas: Effects of the initial substrate and biomass concentrations. Int. J. Hydrog. Energy 2008, 33, 6109–6115. [Google Scholar] [CrossRef]
- Chu, C.-F.; Li, Y.-Y.; Xu, K.-Q.; Ebie, Y.; Inamori, Y.; Kong, H.-N. A pH- and temperature-phased two-stage process for hydrogen and methane production from food waste. Int. J. Hydrog. Energy 2008, 33, 4739–4746. [Google Scholar] [CrossRef]
- Lee, Y.-W.; Chung, J. Bioproduction of hydrogen from food waste by pilot-scale combined hydrogen/methane fermentation. Int. J. Hydrog. Energy 2010, 35, 11746–11755. [Google Scholar] [CrossRef]
- Lay, J.J.; Fan, K.S.; Chang, I.J.; Ku, C.H. Influence of chemical nature of organic wastes on their conversion to hydrogen by heat-shock digested sludge. Int. J. Hydrog. Energy 2003, 28, 1361–1367. [Google Scholar] [CrossRef]
- Kim, S.-H.; Han, S.-K.; Shin, H.-S. Feasibility of biohydrogen production by anaerobic co-digestion of food waste and sewage sludge. Int. J. Hydrog. Energy 2004, 29, 1607–1616. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, S.H.; Kim, H.W.; Kim, M.S.; Shin, H.S. Sewage sludge addition to food waste synergistically enhances hydrogen fermentation performance. Bioresour. Technol. 2011, 102, 8501–8506. [Google Scholar] [CrossRef]
- Dong, L.; Zhenhong, Y.; Yongming, S.; Xiaoying, K.; Yu, Z. Hydrogen production characteristics of the organic fraction of municipal solid wastes by anaerobic mixed culture fermentation. Int. J. Hydrog. Energy 2009, 34, 812–820. [Google Scholar] [CrossRef]
- Koch, K.; Plabst, M.; Schmidt, A.; Helmreich, B.; Drewes, J.E. Co-digestion of food waste in a municipal wastewater treatment plant: Comparison of batch tests and full-scale experiences. Waste Manag. 2016, 47, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Pecorini, I.; Baldi, F.; Albini, E.; Galoppi, G.; Bacchi, D.; Della Valle, A.; Baldi, A.; Bianchini, A.; Figini, A.; Rossi, P.; et al. Hydrogen production from food waste using biochemical hydrogen potential test. Procedia Environ. Sci. Eng. Manag. 2017, 4, 155162. [Google Scholar] [CrossRef]
- Keskin, T.; Arslan, K.; Abubackar, H.N.; Vural, C.; Eroglu, D.; Karaalp, D.; Yanik, J.; Ozdemir, G.; Azbar, N. Determining the effect of trace elements on biohydrogen production from fruit and vegetable wastes. Int. J. Hydrog. Energy 2018, 1–12. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameter | Measure Unit | FW | AS | Test | |||
|---|---|---|---|---|---|---|---|
| ISR 0.05 | ISR 0.08 | ISR 0.14 | ISR 0.25 | ||||
| pH 1 | --- | 5.5 ± 0.2 | 7.1 ± 0.02 | 6.4 ± 0.2 | 6.5 ± 0.1 | 6.7 ± 0.2 | 6.9 ± 0.2 |
| TS | % | 18.8 ± 0.5 | 0.6 ± 0.09 | 8.8 ± 0.02 | 7.0 ± 0.05 | 5.2 ± 0.07 | 3.3 ± 0.08 |
| VS | % TS | 95.6 ± 2.9 | 61.7 ± 4.7 | 94.3 ± 0.06 | 93.7 ± 0.1 | 92.6 ± 0.06 | 90.4 ± 0.05 |
| TOC | % TS | 46.2 ± 0.1 | 36.4 ± 0.1 | 45.8 ± 0.1 | 45.7 ± 0.1 | 45.3 ± 0.1 | 44.7 ± 0.2 |
| TN | % TS | 2.8 ± 0.1 | 6.4 ± 0.2 | 2.9 ± 0.4 | 3.0 ± 0.3 | 3.1 ± 0.3 | 3.4 ± 0.1 |
| sCarb | g/L 2 | ND | ND | 21.8 ± 1.2 | 16.5 ± 0.8 | 12.2 ± 0.5 | 7.8 ± 0.8 |
| Type of Substrate | Type of Inoculum | Inoculum Pre-Treatment | ISR (g VSinoculum/g VSFW) | pH | T (°C) | Reactor Operation Mode | H2 Yield (mL H2/g VSFW) | Reference |
|---|---|---|---|---|---|---|---|---|
| FW | Activated sludge | - | 0.14 | 6.5 | 39 | Batch | 89.8 | Present study |
| FW | Anaerobic sludge | - | 0.23 a | 5.5 b (n.c.) | 36 | Batch | 56.5 | [14] |
| FW | Anaerobic sludge | - | 0.17 | 6.3 b (n.c.) | 35 | Batch | 39 | [19] |
| FW | Anaerobic sludge | - | 0.14 | 6.6 b (n.c.) | 50 | Batch | 57 | [19] |
| FW | Anaerobic sludge | HST c | 0.13 | 6.0 b (n.c.) | 37 | Batch | 55 | [13] |
| FW | Anaerobic sludge | HST | 2 | 4.5 b (n.c.) | 55 | Batch | 60.6 | [10] |
| FW | Anaerobic sludge | HST | 1 | 5.0 | 55 | Batch | 60.3 | [10] |
| Parameter | Measure Unit | Test | |||
|---|---|---|---|---|---|
| ISR 0.05 | ISR 0.08 | ISR 0.14 | ISR 0.25 | ||
| SHPmax | L H2/kg VSFW | 49.8 | 56.6 | 88.8 | 71.0 |
| Rmax | L H2/kg VSFW∙h | 4.7 | 7.9 | 10.8 | 6.8 |
| λ | h | 20.6 | 4.7 | 3.1 | 2.3 |
| t95 | h | 30.8 | 11.8 | 11.1 | 12.4 |
| R2 | - | 0.999 | 0.997 | 0.990 | 0.987 |
| Test | HAc | HPr | HBu | HVal | HHex | HHep | EtOH |
|---|---|---|---|---|---|---|---|
| mmol/gVSFW | |||||||
| ISR 0.05 | 2.51 ± 0.06 | 2.53 ± 0.06 | 0.73 ± 0.03 | 0.10 ± 0.004 | 0.15 ± 0.01 | 0.39 ± 0.01 | 0.06 ± 0.004 |
| ISR 0.08 | 1.80 ± 0.08 | 1.10 ± 0.06 | 1.51 ± 0.03 | 0.09 ± 0.004 | 0.13 ± 0.01 | 0.59 ± 0.02 | 1.00 ± 0.06 |
| ISR 0.14 | 1.82 ± 0.10 | 1.49 ± 0.08 | 1.07 ± 0.03 | 0.17 ± 0.006 | 0.31 ± 0.02 | 1.12 ± 0.03 | 0.06 ± 0.002 |
| ISR 0.25 | 3.46 ± 0.09 | 1.37 ± 0.05 | 1.52 ± 0.05 | 0.07 ± 0.003 | 0.03 ± 0.002 | 0.03 ± 0.001 | 0.79 ± 0.01 |
| Test | DOC | VFAs | EtOH | sCarb | Accounted DOC | Acidification Yield | Conversion Efficiency |
|---|---|---|---|---|---|---|---|
| g C | % | % | mol H2/mol Hexose | ||||
| ISR 0.05 | 75.5 ± 2.3 | 67.9 ± 3.1 | 0.7 ± 0.02 | 1.6 ± 0.04 | 93.0 ± 1.2 | 89.9 ± 1.4 | 0.34 ± 0.004 |
| ISR 0.08 | 55.1 ± 2.1 | 41.8 ± 2.4 | 4.6 ± 0.1 | 1.4 ± 0.1 | 86.9 ± 0.8 | 76.0 ± 1.5 | 0.32 ± 0.01 |
| ISR 0.14 | 44.0 ± 1.8 | 43.5 ± 2.3 | 0.2 ± 0.01 | 1.1 ± 0.04 | 102.0 ± 1.2 | 98.9 ± 1.2 | 0.59 ± 0.003 |
| ISR 0.25 | 20.5 ± 1.2 | 16.9 ± 0.9 | 1.5 ± 0.02 | 0.5 ± 0.02 | 92.0 ± 0.8 | 82.3 ± 0.4 | 0.43 ± 0.01 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappai, G.; De Gioannis, G.; Muntoni, A.; Spiga, D.; Boni, M.R.; Polettini, A.; Pomi, R.; Rossi, A. Biohydrogen Production from Food Waste: Influence of the Inoculum-To-Substrate Ratio. Sustainability 2018, 10, 4506. https://doi.org/10.3390/su10124506
Cappai G, De Gioannis G, Muntoni A, Spiga D, Boni MR, Polettini A, Pomi R, Rossi A. Biohydrogen Production from Food Waste: Influence of the Inoculum-To-Substrate Ratio. Sustainability. 2018; 10(12):4506. https://doi.org/10.3390/su10124506
Chicago/Turabian StyleCappai, Giovanna, Giorgia De Gioannis, Aldo Muntoni, Daniela Spiga, Maria Rosaria Boni, Alessandra Polettini, Raffaella Pomi, and Andreina Rossi. 2018. "Biohydrogen Production from Food Waste: Influence of the Inoculum-To-Substrate Ratio" Sustainability 10, no. 12: 4506. https://doi.org/10.3390/su10124506